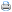Página anterior Página anterior | Voltar ao início do trabalho | Página seguinte  |
O experimento foi realizado no setor de Avicultura da Faculdade de Medicina Veterinária e Zootecnia da UNESP, Campus de Botucatu, SP, no período de abril a maio de 2001. Foram utilizados 2.925 pintos de um dia de idade, sexados e distribuídos em 36 unidades experimentais (UE) de 5 m2 cada, em um galpão coberto com telhas onduladas de fibrocimento, com pé-direito de 3,5 m, largura de 8 m e comprimento de 40 m, equipado com ventiladores e dotado de cortinas laterais. Cada unidade experimental foi equipada com uma campânula com lâmpada de infravermelho de 250W, um bebedouro pendular e dois comedouros tubulares. As temperaturas no interior do galpão variaram de 18,5 a 28,9°C, com média de 23,7°C, em todo o período experimental.
As aves foram distribuídas segundo um delineamento inteiramente casualizado, em arranjo fatorial 3x3x2, ou seja, três linhagens (Ross 308, Cobb 500 e Hybro PG) e três densidades (10, 13 e 16 aves/m2) e dois sexos, com duas repetições e número variável de aves, em função da densidade populacional. A análise estatística dos dados foi realizada por intermédio do procedimento GLM do SAS (SAS, 1998).
Foi utilizado um programa de alimentação com três fases: inicial de 1 a 21 dias, crescimento de 22 a 35 dias e final de 36 a 42 dias (Tabela 1). Para avaliação das características de desempenho, as aves foram pesadas com um dia de idade e, a seguir, ao final de cada fase. Foram avaliados o ganho de peso, o consumo de ração, a conversão alimentar, a mortalidade e as produções em quilos de peso vivo/m2. Além destas características, foi feita uma análise econômica da produção de quilos de peso vivo por 100 m2 de galpão, de acordo com exposto por Luchesi (1998). O cálculo do custo do quilo de ração foi realizado com base nos preços das matérias-primas utilizadas e o preço de kg do frango vivo e do pintinho de um dia foram cotados no mês de dezembro de 2001, no Estado de São Paulo. O custo médio da ração calculado foi de US$ 0,164/kg; o preço do frango vivo, de US$ 0,50/kg; e o preço do pintinho de um dia, de US$ 0,150.
Aos 42 dias de idade, foram retiradas seis aves por parcela, ou seja, 216 no total, para determinação do rendimento de carcaça e partes, bem como para avaliação da qualidade da carne de peito. As aves foram abatidas após jejum de 8 horas em um abatedouro experimental.
Foram avaliadas as seguintes características: peso vivo, peso da carcaça, peso de pernas, peso do peito total, peso da carne do peito, peso do osso do peito, peso da pele do peito, peso de asas, peso de dorso e porcentagem de gordura abdominal. O rendimento de carcaça foi calculado em relação ao peso vivo antes do abate [%RC = (peso carcaça*100/peso vivo)] e o rendimento das partes da carcaça e peito, em função do peso da carcaça [%RP = (peso da parte*100/peso carcaça].
Para determinação da perda de peso por cozimento, foram utilizados os filés inteiros, que, após pesados degelados, foram embalados em papel alumínio e cozidos em uma chapa de aquecimento uniforme até atingir a temperatura interna de 82°C ± 2°C. A seguir, as amostras foram colocadas sobre papel absorvente para resfriamento até atingir a temperatura de 20 a 25°C. Novamente, as amostras foram pesadas e a perda de peso logo após o cozimento, determinada. Estas amostras foram envolvidas com papel absorvente (para remoção da umidade superficial), embaladas em sacos plásticos e mantidas sob refrigeração a 4°C, por 24 horas, para determinação da maciez (Honikel, 1987). As amostras utilizadas para determinação da perda de peso por cozimento também foram usadas para determinação da força de cizalhamento (maciez). Os filés esquerdos foram separados e destes, retiradas amostras na forma de paralelepípedo com 2x2x1 (13 cm), que foram colocadas com as fibras orientadas no sentido transversal às lâminas do aparelho Warner-Bratzler, acoplado ao aparelho Instron M 2318, conforme técnica descrita por Froning et al. (1978).
A determinação do pH foi feita pelo eletrodo de penetração, diretamente no peito das aves, 24 horas post mortem. Essa determinação foi realizada em todas as aves amostradas para avaliação do rendimento de carcaça.
As características de desempenho são apresentadas nas Tabelas de 2 a 6. A mortalidade não foi influenciada pelos fatores utilizados nas fases de criação das aves.
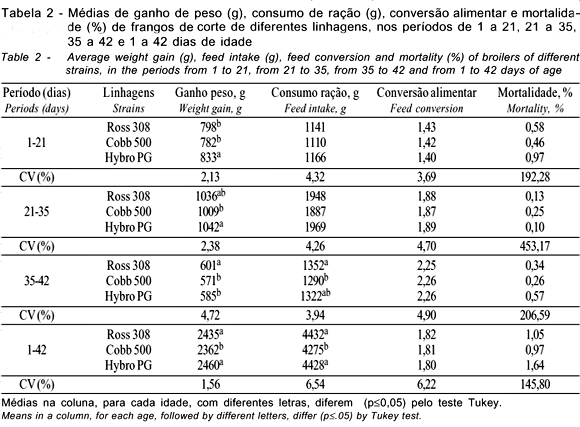

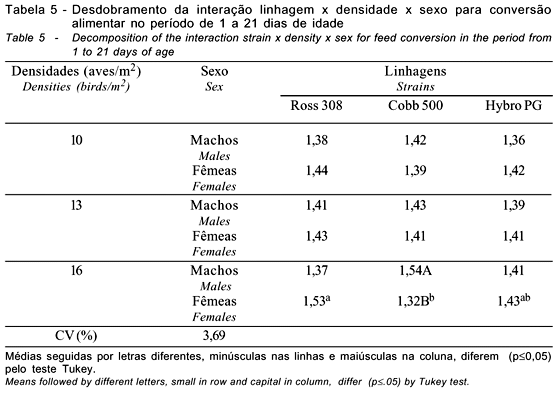
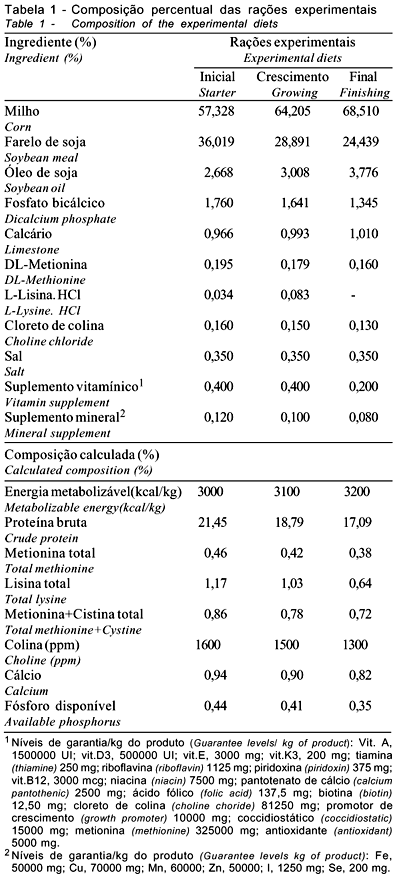
Na fase inicial (1 a 21 dias), houve interação significativa (p<0,05) entre linhagem e sexo para o consumo de ração (Tabela 3). Observa-se que o consumo de ração foi influenciado (p<0,05) pelas linhagens apenas entre as fêmeas e apenas no consumo de ração da linhagem Cobb 500 os machos apresentaram maior valor que as fêmeas. No mesmo período, houve interação significativa entre linhagem x densidade x sexo, havendo diferenças (p<0,05) para a conversão alimentar na densidade de 16 aves/m2, sendo que as linhagens diferiram entre as fêmeas, sendo a linhagem Ross 308 igual à Hybro PG e maior que a Cobb 500, não havendo diferenças entre Cobb 500 e Hybro PG. Apenas na linhagem Cobb 500 houve diferença entre o sexo, sendo que as fêmeas apresentaram melhor conversão alimentar que os machos. Apesar de não haver interação significativa para o ganho de peso, o mesmo foi influenciado (p<0,05) pelas linhagens e densidades testadas, observando-se redução no ganho de peso com o aumento da densidade. Os machos apresentaram melhor ganho de peso que as fêmeas.
Os resultados até os 21 dias indicam diferenças entre as linhagens estudadas e efeitos negativos sobre as características de desempenho, ao se elevar a densidade populacional das aves nesta fase. Estes resultados corroboram os achados de Lisboa et al. (1999), que, avaliando o desempenho de três grupos genéticos obtidos na Universidade Federal de Viçosa, também observaram efeito da linhagem sobre o ganho de peso, o consumo de ração e a conversão alimentar. Lana et al. (1995), Mizubuti et al. (1994), Souza et al. (1994) e Yaissle & Lilburn (1998) também verificaram diferenças para ganho de peso e consumo de ração, ao estudarem o desempenho de diferentes linhagens na fase inicial, porém não observaram efeitos sobre a conversão alimentar. No entanto, Abreu et al. (1996) e Farran et al. (2000), avaliando as mesmas características, não verificaram efeito das linhagens sobre o ganho de peso e consumo de ração, apesar de também terem verificado efeito sobre a conversão alimentar. Nobre et al. (1994) também não observaram efeito das linhagens sobre o ganho de peso e a conversão alimentar, porém, verificaram efeito sobre o consumo de ração. Os resultados obtidos para o ganho de peso na fase inicial, nas diferentes densidades, diferem dos achados de Mizubuti et al. (1994), Stringhini (1998) e Lana et al. (2001b), que não observaram efeito da densidade para esta característica, porém corroboram os achados de Goldflus (1997b), que obteve resultados semelhantes na época de inverno. O consumo de ração verificado confirma os achados de Mizubuti et al. (1994) e Lana et al. (2001a), que também não verificaram influência da densidade sobre esta característica, mas discorda dos resultados encontrados por Goldflus (1994) e Stringhini (1998).
Lana et al. (2001b) e Stringhini (1998) verificaram piora na conversão alimentar com o aumento da densidade, o que corrobora os dados encontrados neste trabalho, embora não tenha havido significância. Goldflus (1994), por sua vez, também não verificou diferença nesta característica na época de inverno.
Na fase de crescimento (21 a 35 dias), pode-se observar que as linhagens apresentaram diferenças apenas para o ganho de peso e as densidades avaliadas não tiveram influência sobre as características avaliadas. O sexo das aves influenciou o ganho de peso, o consumo de ração e a conversão alimentar, sendo os machos superiores às fêmeas em todos eles, exceto na conversão alimentar (Tabela 6). Dados semelhantes quanto às linhagens de frangos de corte foram obtidos por Nobre et al. (1994), Lana et al. (1995) e Lisboa et al. (1999), que também observaram diferenças para ganho de peso na fase de crescimento, porém foi constatado efeito sobre o consumo de ração. Lana et al. (2001a), avaliando as densidades de 10, 12 e 16 aves/m2, também não verificaram efeitos das densidades nos parâmetros de desempenho nesta fase.
Na fase de crescimento (21 a 35 dias), as linhagens apresentaram diferenças apenas para o ganho de peso e as densidades avaliadas não tiveram influência sobre as características avaliadas. O sexo das aves influenciou o ganho de peso, o consumo de ração e a conversão alimentar, sendo os machos superiores às fêmeas, exceto na conversão alimentar (Tabela 6). Dados semelhantes com relação às linhagens de frangos de corte foram obtidos por Nobre et al. (1994), Lana et al. (1995) e Lisboa et al. (1999), que também observaram diferenças para ganho de peso na fase de crescimento, assim como para consumo de ração. Lana et al. (2001a) não verificaram efeitos de densidade (10, 12 e 16 aves/m2) sobre as características de desempenho, nesta fase.
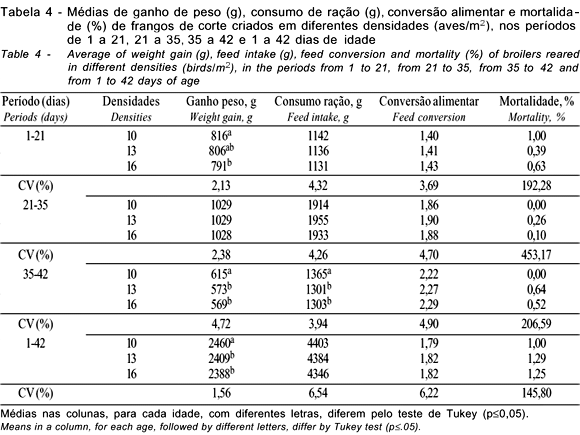
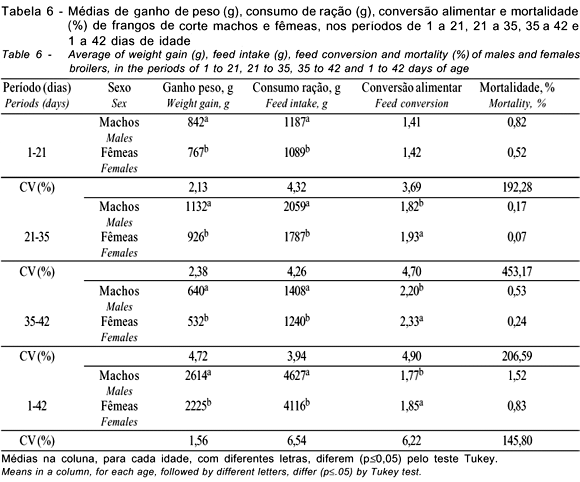
Na fase final de criação (35 a 42 dias), as linhagens e as densidades apresentaram diferenças para ganho de peso e consumo de ração, sendo que a densidade de 10 aves/m2 proporcionou maior consumo de ração e, conseqüentemente, maior ganho de peso, não havendo diferenças entre as densidades de 13 e 16 aves/m2. O sexo também influenciou as características avaliadas, sendo os machos superiores às fêmeas, exceto na conversão alimentar. Estes resultados corroboram os achados de Mizubuti et al. (1994) e Lana et al. (1995), que também encontraram diferenças para o consumo de ração e ganho de peso na fase final de criação de frangos de corte ao avaliarem diferentes linhagens. Os dados para as densidades confirmam os achados de Lana et al. (2001b) que também verificaram um maior consumo de ração para a menor densidade avaliada, porém, estes mesmos autores não observaram efeito sobre o ganho de peso.
No período total de criação (1 a 42 dias), houve diferenças entre as linhagens avaliadas para ganho de peso e consumo de ração. Resultados semelhantes foram encontrados por Mizubuti et al. (1994), Nobre et al. (1994), Lana et al. (1995) e Lisboa et al. (1999), que também observaram diferenças para estas características, ao avaliarem o desempenho de diferentes linhagens comerciais. Sousa et al. (1994), Smith et al. (1998) e Fernandes et al. (2001) encontraram resultados semelhantes, mas também verificaram efeitos das linhagens sobre a conversão alimentar. As densidades influenciaram o ganho de peso (p<0,05); as aves criadas na densidade de 10 aves/m2 apresentaram melhor ganho que aquelas criadas nas densidades de 13 e 16 aves/m2, que não diferiram entre si. Isto se justifica pelo fato destas aves terem apresentado maior consumo de ração e melhor conversão alimentar, embora nestas características não tenham sido encontradas diferenças significativas (p>0,05).
Avaliando o período total de criação, Stringhni et al. (1998) e Moreira et al. (2001) verificaram diferenças para ganho de peso, ao se variar a densidade de criação, porém, Goldflus et al. (1997a) e Moreira et al. (2001) observaram efeito da densidade sobre o consumo de ração. No entanto, Mizubuti et al. (1994) e Lana et al. (2001a) não verificaram efeito da densidade sobre as características de desempenho em frangos de corte. Em estudos sobre densidades, geralmente, quando há variação maior que o intervalo de 10 para 16 aves/m2, as diferenças nas características de desempenho são mais acentuadas. Porém, no intervalo de 10 para 16 aves/m2 estas diferenças são menores e, às vezes, inexistem, sendo que, de 12 a 16 aves/m2, raramente ocorrem diferenças. O sexo das aves influenciou as características de desempenho, sendo os machos superiores às fêmeas em todas elas.
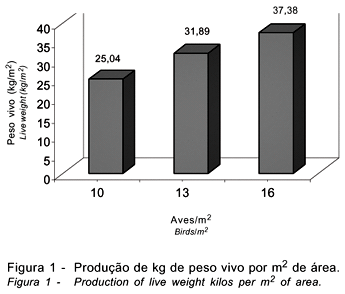
A produção de peso vivo /m2 está apresentada na Figura 1 e a análise econômica, na Tabela 7. À medida que se eleva a densidade de criação, aumenta a produção de peso vivo/m2. Houve incremento de 27,36 e 49,28%, respectivamente, na produção de peso vivo/m2, com o aumento da densidade (de 10 para 13 e 16 aves/m2), resultados que corroboram os achados de Goldflus et al. (1997a) e Stringhini et al. (1998), que observaram maior produção por unidade de área para as aves criadas em alta densidade.
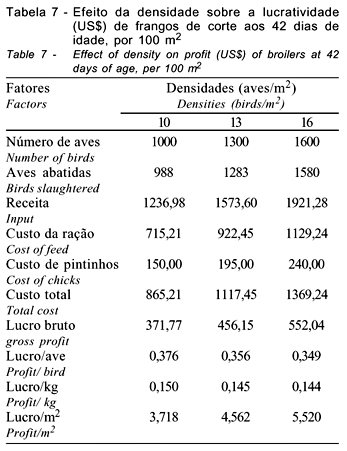
Pode-se observar, na Tabela 7, que a receita e o custo total aumentam, à medida que se eleva a densidade de criação, obtendo-se maior lucro bruto com o aumento da densidade. Todavia, ao se avaliar o lucro por ave e por kg de frango vivo, nota-se que estes diminuem com o aumento da densidade, ao passo que o lucro por m2 cresce, atingindo 22,70 e 48,47% quando se passa de 10 para 13 e 16 aves/m2. Assim, sempre ocorrerá porcentagem de ganho, ao se elevar a densidade, embora isto possa ser maximizado ou minimizado de acordo com o preço das matérias-primas e os custos com readequação de equipamentos e manejo.
As linhagens diferiram (p<0,05) para peso vivo, rendimento de peito, pernas, asas, dorso e gordura abdominal (Tabela 8). Houve interações significativas (p<0,05) entre linhagens x sexo para rendimento de peito, pernas e gordura abdominal, sendo que, para o rendimento de peito, as linhagens diferiram tanto entre os machos como entre as fêmeas; duas das três linhagens avaliadas apresentaram diferenças significativas (p<0,05) entre machos e fêmeas, em que as fêmeas apresentaram maiores rendimentos. Para o rendimento de pernas, apenas os machos diferiram entre si, sendo superiores às fêmeas (p<0,05) nesta característica. Para a gordura abdominal, apenas uma das linhagens apresentou diferenças entre machos e fêmeas, observando-se as maiores porcentagens para as fêmeas. Os machos foram superiores às fêmeas para peso vivo, rendimento de carcaça e pernas e inferiores para rendimento de asas. Embora as linhagens não tenham apresentado diferenças entre o rendimento de carcaça, houve maior especificidade para o rendimento de partes. Mendes et al. (1993) e Fernandes et al. (2001) também não observaram diferenças para o rendimento de carcaça, ao avaliarem diferentes linhagens, porém outros autores (Souza et al., 1994; Smith et al., 1998; Figueiredo et al., 1999; Farran et al., 2000) mencionaram diferenças entre linhagens para rendimento de carcaça. Resultados semelhantes para rendimento de peito foram verificados por Lisboa et al. (1999), Figueiredo et al. (1999), Araújo et al. (1999) e Fernandes et al. (2001), que também observaram diferenças nesta característica ao estudarem diferentes linhagens, comprovando que os programas de melhoramento adotados pelas empresas têm resultados bastante diferenciados para esta característica.
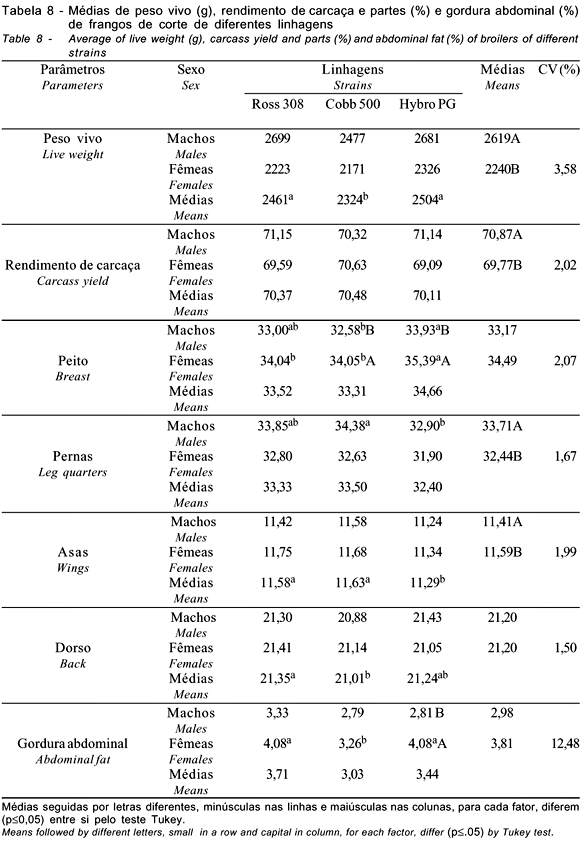
Mendes et al. (1993), Souza et al. (1994) e Abreu et al. (1996) não verificaram efeito das linhagens para rendimento de asas, resultados que corroboram os achados de Araújo et al. (1999), que obtiveram resultados semelhantes em diferentes linhagens. Os resultados obtidos para porcentagem de gordura abdominal estão de acordo com relatos de Abreu et al. (1996), Figueiredo et al. (1999), Araújo et al. (1999) e Farran et al. (2000). Outros autores (Souza et al., 1994; Lisboa et al., 1999; Fernandes et al., 2001), porém, não verificaram diferenças na porcentagem de gordura abdominal, em diferentes linhagens comerciais.
Observa-se, na Tabela 9, que as densidades não apresentaram diferenças para as características avaliadas. Para o rendimento de pernas, houve diferenças entre machos e fêmeas nas densidades de 10 e 13 aves/m2, sendo os machos superiores às fêmeas. Esperava-se o mesmo efeito em todas as densidades, pois os machos, geralmente, mostram-se superiores às fêmeas para esta característica. Portanto, não há justificativa para ausência de diferença na densidade de 16 aves/m2. A porcentagem de gordura abdominal foi diferente entre machos e fêmeas, nas densidades de 10 e 16 aves/m2, observando-se as maiores porcentagens para as fêmeas. A ausência de diferença na densidade de 13 aves/m2 não permite inferência sobre este dado, pois, geralmente, as fêmeas têm maiores teores de gordura abdominal que os machos.
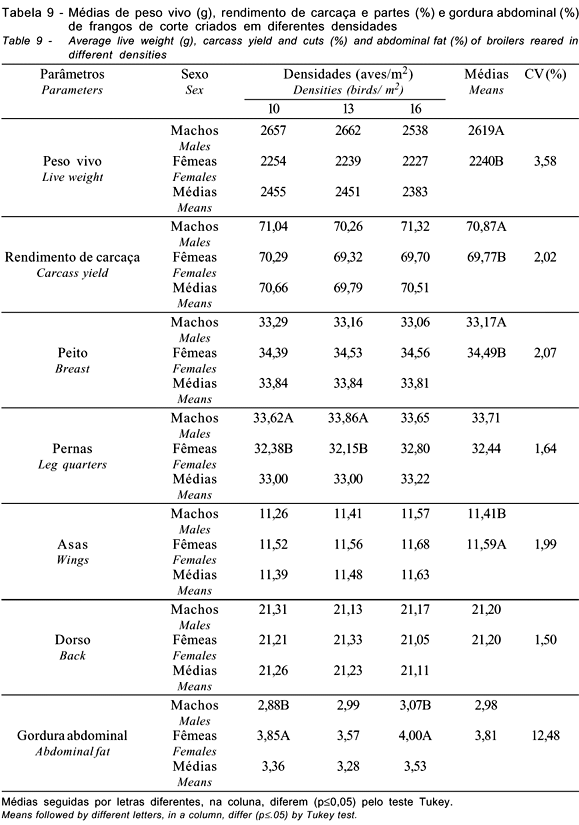
Embora não tenham sido observadas diferenças significativas (p>0,05), nota-se tendência de aumento da gordura abdominal, com a elevação da densidade de criação. Contudo, os resultados obtidos neste trabalho indicam que a variação da densidade de criação de 10 a 16 aves/m2 não afeta o rendimento de carcaça e partes. Estes resultados corroboram os achados de Stringhini et al. (1997), avaliando densidades de 12 e 18 aves/m2, e de Lana et al. (2001), estudando densidades de 10, 12 e 16 aves/m2. Goldflus et al. (1997a), Stringhini et al. (1998) e Campos et al. (1999), utilizando variação da densidade maior que a deste trabalho, apesar de não terem observado diferença para o rendimento de partes e gordura abdominal, verificaram efeito sobre o rendimento de carcaça. Goldflus et al. (1997b), ao variarem as densidades de 10 a 22 aves/m2, também observaram diferenças para rendimento de carcaça, apenas no período de inverno. Moreira et al. (2001), avaliando as densidades de 10, 13 e 16 aves/m2, não verificaram diferenças para o rendimento de carcaça e das principais partes (peito e pernas), mas observaram influência sobre o rendimento de patas e gordura abdominal.
Na Tabela 10, são apresentados os resultados de cortes nobres (peito e pernas). As linhagens avaliadas apresentaram diferenças para carne de peito, osso de peito, osso de pernas e pele de pernas, enquanto as densidades avaliadas diferiram apenas para o osso de pernas, comprovando que, com o aumento da densidade, obtém-se maior peso do osso das pernas. O sexo não afetou a porcentagem de osso do peito, sendo os machos superiores às fêmeas para carne e osso de pernas e inferiores para carne do peito e pele do peito e das pernas. Smith et al. (1998) e Souza et al. (1994) também encontraram diferenças para rendimento da carne do peito, ao avaliarem diferentes linhagens. Souza et al. (1994) observaram o efeito das linhagens sobre os rendimentos da carne de peito e de osso do peito nas fêmeas, mas não sobre a pele do peito. Mendes et al. (1993) não verificaram efeito das linhagens para carne, osso e pele do peito, mas observaram influência sobre o rendimento de carne de pernas, resultados que diferem dos obtidos neste trabalho. Mendes et al. (2001a), avaliando as densidades de 10, 13 e 16 aves/m2, também não verificaram efeito (p>0,05) da densidade sobre o rendimento de carne, osso e pele do peito, o que corrobora os achados deste trabalho. O rendimento de filé do peito e os parâmetros de qualidade da carne do peito são apresentados na Tabela 11. Pode-se observar que as linhagens apresentaram diferenças para o peso dos filés e para perda de peso por cozimento, em decorrência das diferenças entre as linhagens para o rendimento de peito e carne de peito, discutidos anteriormente. Souza et al. (1994) também verificaram diferenças no peso dos filés de peito, ao avaliarem diferentes linhagens.
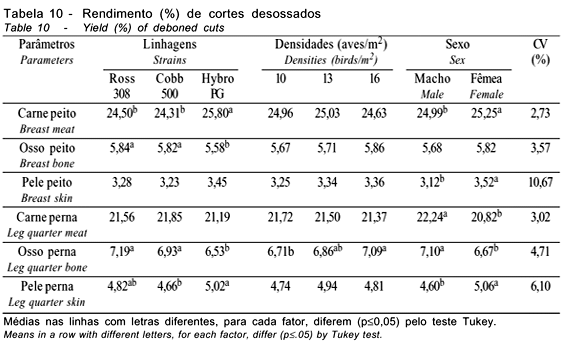
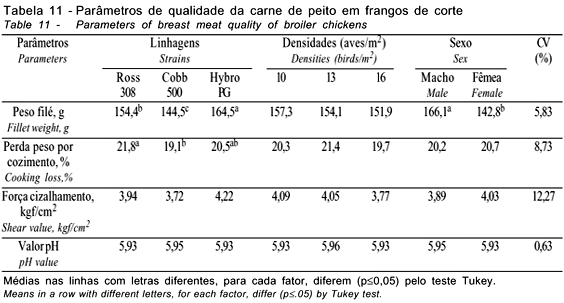
As densidades não influenciaram (p>0,05) o peso dos filés e as características de qualidade avaliadas, corroborando os achados de Mendes et al. (2001b), que também não verificaram influência sobre estas características, trabalhando com as mesmas densidades de criação.
O sexo teve influência apenas sobre o peso dos filés, observando-se o maior peso para os machos. Isto se justifica pelo fato de os mesmos terem maior peso e, conseqüentemente, peitos mais pesados e maiores, embora tenham menor rendimento percentual que as fêmeas.
O aumento da densidade populacional de 10 a 16 aves/m2 causa redução no ganho de peso, principalmente na fase final de criação, apesar de não haver diferenças entre 13 e 16 aves/m2. Com o aumento da densidade, há maior produção de quilos de peso vivo/m2, o que possibilita incrementos na renda bruta da criação. Este aumento da densidade não afeta o rendimento de carcaça e cortes nobres (peito e pernas), além de não influenciar as características de qualidade (perda de peso por cozimento, força de cizalhamento (maciez) e pH) da carne de peito.
As linhagens comerciais avaliadas, apesar de não apresentarem diferenças para o rendimento de carcaça, diferiram quanto ao desempenho e o rendimento de partes, principalmente peito, e para perda de peso por cozimento avaliada no filé do peito.
Os machos apresentaram melhores desempenho, rendimento de carcaça e de partes, porém não diferiram das fêmeas nas características de qualidade da carne.
ABREU, V.M.N.; SILVA, M.A.; TORRES, R.A. et al. Características produtivas de linhagens de corte. Revista Brasileira de Zootecnia, v.25, n.1, p.83-91, 1996.
ARAÚJO, L.F.; JUNQUEIRA, O.M.; ARAÚJO, C.S.S. et al. Diferentes níveis de lisina para duas linhagens de frangos de corte na fase final de criação. Revista Brasileira de Ciência Avícolas, v.1, p.43, 1999. (supl.)
BERAQUET, N. Influência de fatores ante e post mortem na qualidade da carne de aves. Revista Brasileira de Ciência Avícolas, v.1, p.155-166, 1999.
CAMPOS, S.S. Efeitos da energia dietética, densidade populacional, altura de cama e época do ano sobre parâmetros termorreguladores, zootécnicos e da cama em frangos de corte. Jaboticabal: Universidade Estadual Paulista, 1999. 62p. Tese (Doutorado em Zootecnia) - Universidade Estadual Paulista, 1999.
CASTILLO, C.J.C. Qualidade de carcaça e carne de aves. In: CONGRESSO BRASILEIRO DE CIÊNCIA E TECNOLOGIA DE CARNES, 1., 2001, São Pedro. Anais... São Pedro: Instituto de Tecnologia de Alimentos, 2001. p.79-99.
FARRAN, M.T.; KHALIL, R.F.; UWAYJAN, M.G. et al. Performance and carcass quality of commercial broiler strain. Journal of Applied Poultry Research, v.9, n.2, p.252-257, 2000.
FERNANDES, L.M.; VIEIRA, S.L.; KINDLEIN, G. et al. Avaliação do crescimento e rendimento de carcaça de linhagens comerciais e dois tipos de bebedouro. Revista Brasileira de Ciência Avícolas, v.3, p.1, 2001. (Suplemento)
FIGUEIREDO, E.A.P.; ROSA, P.S.; SCHEUERMANN, G.N. et al. Estudos de características de carcaça em frangos de corte. Revista Brasileira de Ciência Avícolas, v.1, p.62, 1999. (Suplemento)
GOLDFLUS, F. Viabilidade da criação de frangos de corte sob alta densidade populacional. Jaboticabal: Universidade Estadual Paulista, 1994. 126p. Dissertação (Mestrado em Zootecnia) - Universidade Estadual Paulista, 1994.
FRONING, G.W.; BABJI, A.S.; MATHER, F.B. The effect of preslaughter temperatures, stress, atruggle and anesthetization on color and textural characteristics of turkey muscle. Poultry Science, v.57, n.3, p.630-633, 1978.
GOLDFLUS, F.; ARIKI, J.; KRONKA, S.N. et al. Efeitos da densidade populacional e da energia da dieta sobre o desempenho de frangos de corte. Revista Brasileira de Zootecnia, v.26, n.2, p.310-315, 1997a.
GOLDFLUS, F.; ARIKI, J.; KRONKA, S.N. et al. Efeitos de diferentes densidades populacionais nas estações fria e quente do ano sobre o desempenho de frangos de corte. Revista Brasileira de Zootecnia, v.26, n.5, p.948-954, 1997b.
HELLMEISTER, P.; CUSTÓDIO, R.W.S.; COELHO, A.A.D. et al. Broiler performance when raised in different floor space densities. Revista Brasileira de Zootecnia, v.27, n.1, p.137-142, 1998.
HONIKEL, K.O. The water binding of meat. Fleischwirttssch, v.67, p.1098-1102, 1987.
LANA, G.R.Q.; SILVA, D.J.; SILVA, M.A. et al. Desempenho comparativo de marcas comerciais e de cruzamentos de diferentes linhagens de frangos de corte produzidos na UFV, em diferentes níveis de energia. 1. Consumo de ração, ganho de peso e conversão alimentar. Revista Brasileira de Zootecnia, v.24, n.5, p.759-767, 1995.
LANA, G.R.Q.; SILVA, R.G.C.; VALERIO, S.R. et al. Estudo técnico-econômico da criação de frangos de corte alojados sob diferentes densidades e programas de alimentação. In: REUNIÃO ANUAL DA SOCIEDADE BRASILEIRA DE ZOOTECNIA, 2001, Piracicaba. Anais... Piracicaba: Sociedade Brasileira de Zootecnia, 2001a. p.712-714.
LANA, G.R.Q.; SILVA, R.G.C.; VALERIO, S.R. et al. Efeito da densidade e de programas de alimentação sobre o desempenho de frangos de corte. Revista Brasileira de Zootecnia, v.30, n.4, p.1258-1265, 2001b. www.scielo.br
LISBOA, J.S.; SILVA, D.J.; SILVA, M.A. et al. Desempenho de três grupos genéticos de frangos de corte alimentados com rações contendo diferentes teores de proteína. Revista Brasileira de Zootecnia, v.28, n.3, p.555-559, 1999.
LUCHESI, J.B. Custo-benefício da criação de frangos de corte em alta densidade no inverno e no verão. In: CONFERÊNCIA APINCO DE CIÊNCIA E TECNOLOGIA AVÍCOLAS, 1998. Campinas. Anais... Campinas: FACTA, 1998. v.3, p.241-248.
MENDES, A.A.; GARCIA, E.A.; GONZALES, E. et al. Efeito da linhagem e idade de abate sobre o rendimento de carcaça de frangos de corte. Revista Brasileira de Zootecnia, v.22, n.3, p.466-472, 1993.
MENDES, A.A. Rendimento e qualidade da carcaça de frangos de corte. In: CONFERÊNCIA APINCO DE CIÊNCIA E TECNOLOGIA AVÍCOLAS, 2001, Campinas. Anais... Campinas: FACTA, 2001a. v.3, p.79-99.
MENDES, A.A.; MOREIRA, J.; GARCIA, R.G. et al. Avaliação do rendimento e qualidade da carne de peito em frangos de corte criados com diferentes densidades e níveis de energia na dieta. Revista Brasileira de Ciência Avícolas, v.3, p.38, 2001b. (Suplemento)
MIZUBUTI, I.Y.; FONSECA, N.A.N.; PINHEIRO, J.W. Desempenho de duas linhagens comerciais de frango de corte, criadas sob diferentes densidades populacionais e diferentes tipos de cama. Revista Brasileira de Zootecnia, v.23, n.3, p.476-484, 1994.
MOREIRA, J.; MENDES, A.A.; GARCIA, R.G. et al. Efeito da densidade de criação e do nível de energia da dieta sobre o desempenho e rendimento de carcaça em frangos de corte. Revista Brasileira de Ciência Avícolas, v.3, p.39, 2001. (Suplemento)
NOBRE, R.T.R.; SILVA, D.J.;, TAFURI, M.L. et al. Efeito do nível de energia sobre o desempenho de diferentes grupos genéticos de frangos de corte. Revista Brasileira de Zootecnia, v.23, n.4, p.595-602, 1994.
PERDOMO, C.C. Controle do ambiente e produtividade de frangos de corte. In: REUNIÃO ANUAL DA SOCIEDADE BRASILEIRA DE ZOOTECNIA, 2001, Piracicaba. Anais... Piracicaba: Sociedade Brasileira de Zootecnia, 2001. p.91-110.
STATISTICAL ANALYSES SYSTEM - SAS. Language guides for personal computer. 3.ed. v.12. Cary: 1998. 37p.
SMITH, E.R.; PESTI, G.M.; BAKALLI, R.I. et al. Further studies on the influence of genotype and dietary protein on the performance of briolers. Poultry Science, v.77, n.11, p.1678-1687, 1998. www.bireme.br
SOUZA, P.A.; SOUZA, H.B.A.; CAMPOS, F.P. et al. Desempenho e características de carcaça de diferentes linhagens comerciais de frangos de corte. Revista Brasileira de Zootecnia, v.23, n.5, p.783-791, 1994.
STRINGHINI, J.H.; ARIKI, J.; CAFÉ, M.B. et al. Níveis de proteína para frangos de corte criados em duas densidades populacionais. II. Características de carcaça. In: CONFERÊNCIA APINCO DE CIÊNCIA E TECNOLOGIA AVÍCOLAS, 1997, Campinas. Anais... Campinas: FACTA, 1997. p.24.
STRINGHINI, J.H.; ARIKI, J.; CAFÉ, M.B. et al. Níveis de metionina+cistina para frangos de corte criados em duas densidades populacionais. I. Desempenho. In: CONFERÊNCIA APINCO DE CIÊNCIA E TECNOLOGIA AVÍCOLAS, 1998, Campinas. Anais... Campinas: FACTA, 1998. p.21.
YAISSLE, J.; LILBURN, M.S. Effects of dietary protein and strain on the growth of broiler breeder pullets from zero to five weeks of age. Poultry Science, v.77, n.11, p.1613-1619, 1998. www.bireme.br
1 Parte da tese de Doutorado do primeiro autor.
Joerley MoreiraI; Ariel Antônio MendesII; Roberto de Oliveira RoçaIII; Edivaldo Antônio GarciaII; Irenilza de Alencar NaasIV; Rodrigo Garófallo GarciaI; Ibiara Correia Lima de Almeida PazI - arielmendes[arroba]fca.unesp.br
IZootecnista, Pós-Graduando do Programa de Pós-Graduação da FMVZ-UNESP, Campus de Botucatu - SP. IIProfessores do DPEA da FMVZ-UNESP, Campus de Botucatu - SP IIIProfessor da FCA-UNESP, Campus de Botucatu - SP IVProfessor da FEAGRI-UNICAMP, Campinas SP.
| Página anterior | Voltar ao início do trabalho | Página seguinte |
|
|
|