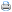Página anterior Página anterior | Voltar ao início do trabalho | Página seguinte  |
Foram utilizados frutos de tomate (Lycopersicon esculentum Mill.), cultivar Kadá, oriundos de plantas não transformadas e transformadas geneticamente com um clone de ADN, denominado pMEL1 (Balagué et alii, 1993), correspondente à ACC oxidase de melão, em orientação antisenso. Os tomateiros foram cultivados em casa de vegetação, localizada no Campus da Universidade Federal de Pelotas - Capão do Leão - RS, no período de outubro de 1994 a março de 1995.
O amadurecimento foi acompanhado em função da evolução da coloração, determinando-se o ciclo total de maturação, a produção de etileno, a síntese da enzima ACC oxidase e a composição físico-química dos frutos.
Escala de coloração
A avaliação da coloração foi realizada utilizando-se uma escala previamente estabelecida por Pratt & Workman (1962), que classifica os frutos de tomate em:
a) verde-imaturos: quando estão em fase de crescimento;
b) verde-maduros: quando adquirem o tamanho máximo e a capacidade de amadurecer;
c) "breaker": quando houver pigmentação amareloavermelhada em até 20% da superfície;
d) vermelhos: quando apresentarem-se completamente avermelhados;
e) senescentes: quando apresentam amolecimento avançado.
Produção de etileno
O etileno sintetizado pelos frutos de tomate foi dosado por cromatografia em fase gasosa, utilizando-se cromatógrafo a gás VARIAN 3300, de acordo com as recomendações descritas por Dupille et alii (1992). As amostras, contendo aproximadamente 1 Kg de frutos, foram acondicionadas, durante 1 hora, em frascos com capacidade de 5 litros, hermeticamente fechados após o que, coletou-se 1 ml da atmosfera gasosa para a dosagem do etileno. A produção deste regulador de crescimento foi expressa em nanolitros de etileno / hora / grama de matéria fresca (nl.h-1.g-1 ).
"Western blot"
O "western blot" foi realizado a partir da metodologia descrita por Meyer et alii (1985), que compreende 4 etapas principais: 1) extração das proteínas; 2) separação eletroforética das proteínas em condições desnaturantes; 3) eletrotransferência das proteínas para membranas de nitrocelulose; 4) reações imunoquímicas de dectecção da ACC oxidase.
Para a extração de proteínas totais, 5 gramas de tomate foram homogeneizados com uma solução de ácido tricloro acético a 15% (p/v), contendo 5% (v/v) de β-mercaptoetanol. Apσs 1 hora de incubação do material a 4oC, a mistura foi centrifugada a 20.000 g durante 30 minutos. O precipitado resultante foi lavado 5 vezes com uma solução de acetato de amônio 100 mM a 20oC dissolvido em metanol. Finalmente, o precipitado foi dissolvido no tampão Laemmlli (Tris-HCl 25 mM, pH 6,5 contendo glicerol a 10%, p/v; β-mercaptoetanol a 5%, v/v; dodecil sulfato de sódio -SDS-a 2,3 %, p/v; e azul de bromofenol a 0,005%, p/v), de maneira a obterse aproximadamente 1µg de proteína. µl-1.de tampão. A dosagem do teor de proteínas foi determinada segundo metodologia descrita por Harlow & Lane (1988).
A separação eletroforética das proteínas foi efetuada em condições desnaturantes, utilizando, para cada amostra, 10 µg de proteínas, em gel de poliacrilamida descontínuo.
O gel de concentração foi constituído de 4% (p/v) de N,N-metilenobisacrilamida, 0,1% (p/v) de SDS, 0,04% (v/v) de Temed, 0,03% (p/v) de persulfato de amônio dissolvidos num tampão Tris-HCl 125 mM, pH 6,8. Já o gel de separação, caracterizou-se por uma maior concentração de N,N-metilenobisacrilamida (12%) e de Tris-HCl (375 mM), pH 8,9.
A migração eletroforética das proteínas foi feita a 75V durante 30 minutos, aumentando-se a voltagem, em seguida, para 150V até o final do processo. O tampão de migração utilizado foi Tris 25 mM - glicina 192 mM (pH 8,3), contendo 0,01% (v/v) de SDS. A migração foi interrompida quando do início da saída do azul de bromofenol do gel de separação.
A eletrotransferência das proteínas para membranas de nitrocelulose 45 µm foi realizada durante 1,5 horas a 150V, num tampão Tris 25 mM -Glicina 192 mM (pH 8,3), contendo 20% (v/v) de metanol. Concluída esta etapa, as membranas foram submetidas às reações imunoquímicas, como segue: lavagem em água deionizada durante 20 minutos; incubação em tampão Tris-HCl 25 mM, pH 7,4, contendo 350 mM de NaCl, 0,01% (p/v) de NaN3 (TBS), 0,05% (v/v) de Tween 20 (TTBS) e 5% (p/v) de leite em pó desnatado, durante 1,5 horas; enxaguagem em TTBS; incubação neste mesmo tampão contendo 2% de leite em pó desnatado e os anticorpos anti-ACC oxidase (Dupille et alii, 1993) diluídos na proporção 1:10.000 (anticorpo : tampão), durante 2 horas; lavagem em TTBS; incubação com o anticorpo segundário anti-coelho, acoplado à fosfatase alcalina (Dako Laboratories), diluído 1:2.000 num tampão TTBS-2% de leite em pó desnatado, durante 1 hora e, finalmente, lavagem em TTBS e revelação utilizando o 5-bromo-4-cloro-3-indol fosfato e o nitro azul tetrazólio (BCIP/NBT-Vector Laboratories) como substratos.
Análises físico-químicas e de rendimento industrial
Os tomates colhidos em cada estágio de maturação foram submetidos, após descascamento, às análises de pH, acidez total, sólidos solúveis, acúcares redutores, sólidos insolúveis em etanol e matéria seca, seguindo-se as recomendações descritas por Goitia et alii (l995).
Análise estatística
O experimento foi realizado segundo um delineamento inteiramente casualizado, com cinco repetições. A comparação de médias foi efetuada através do teste de Duncan ao nível de 5% de probabilidade.
A produção de etileno e o ciclo de maturação dos frutos de tomate, não transformados e transformados, estão representados na Figura 1. Houve um constante incremento na produção de etileno quando os frutos de tomate não transformados passaram do estado verde-imaturo (2 nl.h-1.g-1) e verde-maduro (2 nl.h-1.g-1) para os estágios mais avançados, "breaker" (18 nl.h-1.g1) e vermelhos (22 nl.h-1.g-1) (Figura 1A). Slater et alii (1985) demonstraram que, durante estes estágios de maturação, há a indução de mais de 300 ARNm. Dentre estes, não mais de uma dezena foram caracterizados (Kende, 1993), destacande-se os correspondentes às enzimas celulase, poligalacturonase, pectinametilesterase, ACC sintetase e ACC oxidase (Fernadez-Maculet & Yang, 1992; Grierson et alii, 986; Kende, 1993, Slater et alii, 1985). A indução da síntese destas duas últimas enzimas explicaria o aumento da produção do etileno (Kende, 1993).
Quando os frutos atingiram a fase de senescência, caracterizado pelo amolecimento generalizado dos tecidos, houve uma redução marcante na produção de etileno, atingindo valores da ordem de 7 nl.h-1.g-1. Este comportamento fisiológico é característico na maioria dos produtos climatéricos (Kende, 1993).
Entretanto, quando se analisa a produção de etileno dos tomates transformados, qualquer que seja o estágio de maturação, os valores são extremamente baixos, não ultrapassando 0.5 nl.h-1.g-1. Esta produção de etileno é similar àquela verificada em tecidos com comportamento não climatérico (frutos não climatéricos, folhas, pecíolos, embriões, etc...) e não injuriados (Abeles et alii, 1992; Adams e Yang, 1979; Kende, 1989). Estes resultados indicam que a expressão do clone de ADN pMEL 1 de melão, introduzido em orientação antisenso em plantas de tomate, é capaz de atenuar mais que 90% da produção de etileno dos frutos. O ciclo de maturação, entretanto, não foi afetado pela transformação (Figura 1B). A passagem do estágio verde-maduro para o sensescente demandou 30 dias e 27 dias para os tomates não transformados e transformados, respectivamente. Resultados similares foram obtidos por Ayub (1995), Hamilton et alii (1990) e Schuch et alii (1991).
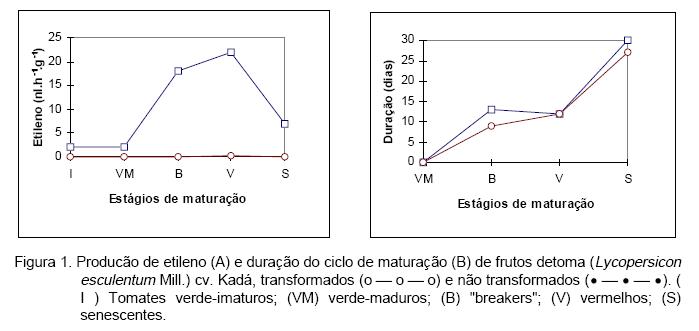
Na Figura 2, está representado o perfil, em "western blot", da proteína ACC oxidase. Semelhantemente à produção do etileno, há um aumento marcante na quantidade desta enzima durante a maturação dos frutos de tomate não transformados. Ressalta-se, no entanto, a presença da ACC oxidase em frutos verde-imaturos (Figura 2A), que se caracterizam pela ausência de gel locular e pela incapacidade de amadurecer após a colheita. Este resultado é coerente com o fato de que todos os tecidos vegetais produzem etileno (Adams & Yang, 1979), mas discordante com os resultados obtidos por Hamilton et alii (1990) que indicam uma ausência total de ARNm correspondentes à ACC oxidase em frutos de tomate em estágios anteriores ao verde-maduro. Esta controvérsia pode ser explicada por uma diferença de sensibilidade entre as técnicas de "western blot" e de "northern blot". É possível, também, que durante o processo evolutivo haja a expressão de diferentes isoformas da ACC oxidase, detectáveis pelos anticorpos policlonais anti-ACC oxidase, mas que não tenham homologia de sequência suficientemente alta para serem postas em evidência por hibridação com a sonda utilizada nos testes moleculares (Rombaldi et alii, 1994).
Quando os tomates não transformados atingem a fase de senescência, há uma redução na produção de etileno (Figura 1A), mas o nível da proteína ACC oxidase mantém-se elevado (Figura 2A). A redução da produção deste regulador de crescimento parece não estar associada à deficiência de substrato da ACC oxidase, pois foi verificado que o aporte de ACC ao meio não restaura a atividade enzimática (Brecht, 1987). Isto indica que a ACC oxidase perde a atividade enzimática durante a degradação-senescência celular, mas preserva sua sequência aminoacídica, já que os níveis desta proteína continuam elevados e o peso molecular conservado.
Em frutos de tomate transformados, apenas pequenas quantidades de proteína ACC oxidase foram postas em evidência (Figura 2B), indicando a eficiência da técnica de transformação na inibição da expressão desta enzima. Estes resultados indicam que a atenuação da produção de etileno deste material se faz pela inibição da síntese da ACC oxidase.
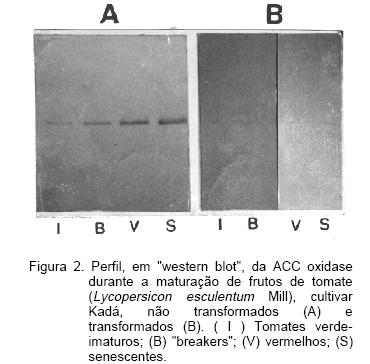
Pela análise dos resultados apresentados na Tabela l pode-se verificar que, para os dois genótipos estudados, afora a acidez total, as demais características de controle de qualidade analisados, não apresentaram variações significativas. Os valores de pH obtidos, situados entre 3,76 e 3,96, são considerados adequados, tanto para o consumo in natura quanto para o processamento industrial dos tomates. Do ponto de vista tecnológico, a faixa de pH mais adequada para o processamento fica entre 3,5 e 4,3 (Goitia et alii, 1995).
Os sólidos solúveis apresentaram-se como os constituintes majoritários da matéria seca dos frutos, não havendo diferenças significativas entre os tomates transformados e não transfomados. Eles representam, em média, 4,27% do peso dos frutos e cerca de 70% dos sólidos totais ou matéria seca. Associados aos teores de sólidos insolúveis em álcool e à renda de descascamento, estes dados permitem inferir sobre os rendimentos industriais durante o processamento de tomate. Assim, após eliminada a casca, que representa, em peso, cerca de 10% dos frutos, os fatores intrínsecos ao produto que determinam o rendimento industrial e as características reológicas, são os teores de sólidos solúveis, de matéria seca e de sólidos insolúveis em álcool. Goitia et alii (1995) citam, por exemplo, que frutos apresentando uma relação SIA (sólidos insolúveis em álcool)/ MS (matéria seca) x l00 superior a 30, conferem aos frutos boas características reológicas e/ou permitem obter subprodutos pastosos de alta consistência. Os frutos aqui analisados apresentaram uma relação SIA/MS x 100 de 32.
A acidez total, apesar de ser mais elevada em frutos de tomate transformados, ainda permanece dentro dos valores médios citados na literatura (Schuch et alii,1991; Goitia et alii,1995). Esta maior acidez dos frutos transformados se deve, provavelmente, a uma menor intensidade respiratória des frutos.
Os resultados permitem concluir que o aumento da produção de etileno durante a crise climatérica de frutos de tomates se faz acompanhada da síntese da enzima ACC oxidase.
A transformação de plantas de tomate com o clone de ADN pMEL l, em orientação antisenso, permite inibir a síntese da ACC oxidase e, por consequência, a do etileno.
Uma redução da produção de etileno em 90% não é suficiente para prolongar o ciclo de maturação de tomates, quando mantidos na planta.
A composição físico-química básica não é afetada pela redução da produção de etileno.
À Fundação de Amparo a Pesquisa do Rio Grande do Sul (FAPERGS) e ao Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) pelo financiamento deste trabalho.
ABELES F. B., MORGAN P. W., SALTVEIT M. E. Jr. Ethylene in plant biology. San Diego. Academic Press, 1992, 414 pp. 2 ed.
ADAMS D. O., YANG S. F. Ethylene biosynthesis: identification of 1-aminocyclopropane-1-carboxylic acid as an intermediate in the conversion of methionine to ethylene. Proc. Natl. Acad. Sci. USA, n.76, p170-174, 1979.
AYUB, R. Transformation génétique du melon. Toulouse: ENSAT, 1995, 135p. Tese (Doutorado em Biologia Celular e Molecular Vegetal), Institut National Polytechnique de Toulouse, 1995.
BALAGUÉ C., WATSON C. F., TURNER . J., ROUGÉ P., PICTON S., PECH J.-C., GRIERSON D. Isolation of a ripening wound-induced cDNA from Cucumis melo L. encoding a protein with homology to the ethylene-forming enzyme. Eur. J. Biochem., n. 276 p27-34, 1993.
BRECHT J.K. Locular gel formation in developing tomato fruit and the initiation of ethylene production. HortScience, n.22, p476-479, 1987.
DUPILLE E., ROMBALDI C. V., LELIÈVRE J. M., CLEYET-MAREL J. C., PECH J. C., LATCHÉ A. Purification, properties and partial amino-acid sequence of 1-aminocyclopropane-1-carboxylic acid oxidase from apple fruits. Planta, n.190, p65-70, 1993.
FERNANDEZ-MACULET J. C. & YANG S. F. Extration and partial characterization of the ethylene-forming enzyme from apple fruit. Plant Physiol., n.99, p751754, 1992.
GOITIA, A.E., QUESTA, A.G.; RANA, E.; RODRIGUEZ,
S. C. Nuevos cultivares de tomate: su aptitud para uso industrial. La alimentacion Latinoamericana, n.207, p26-31, 1995.
GRIERSON D., TUCKER G.A., KEEN J., RAY J., BIRD C.R., SCHUCH W. Sequencing and identification of a cDNA clone for tomato polygalacturonase. Nucl. Acid. Res., n. 14, p85958603, 1986.
HAMILTON A. J., LYCETT G. W., GRIERSON D. Antisens gene that inhibits synthesis of the hormone ethylene in transgenic plants. Nature, n. 346, p284287, 1990.
HAMILTON A. J., BOUZAYEN M., GRIERSON D. Identification of a tomato gene for the ethylene forming enzyme by expression in yeast. Proc. Natl. Acad. Sci. USA, n. 88, p7434-7437, 1991.
HARLOW E., LANE D. Antibodies - A laboratory manual, Cold Spring Harbor Laboratory, Cold Spring Harbor, N.Y., 1988, 689p.
KENDE H. Enzymes in ethylene biosynthesis. Plant Physiol., n. 91, p1-4, 1989.
KENDE H. Ethylene biosynthesis. Annu. Rev. Plant Physiol. Plant Mol. Biol., n.44, p283-307, 1993.
MCGARVEY D. J. & CHRISTOFFERSEN R. E.. Characterization and kinetic parameters of ethyleneforming enzyme from avocado fruit. J. Biol. Chem., n. 267, p5964-5967, 1992.
MEYER Y., GROSSET J., CHARTIER Y., CLEYETMAREL J.C. Preparation by two-dimensional electrophoresis of proteins for antibody production: Antibodies against proteins whose synthesis is reduced by auxin in tobacco mesophyl protoplasts. Electrophoresis, n. 9, p704-712, 1988.
PECH J.C., LATCHE A., BALAGUE C. Cellular and molecular aspects of the plant hormone ethylene. Kluwer publisher. NL., 1993, 385p.
PECH J.C., BALAGUE C., LATCHE A., BOUZAYEN M. Postharvest physiology of climacteric fruits : recent developments in the biosynthesis and action of ethylene. Sci. Alim., n. 14, p3-15, 1994.
PIRRUNG M.C., KAISER L.M., CHEN J. Purification and properties of the apple fruit ethylene-forming enzyme. Biochemistry, n. 32, p7445-7450, 1993.
PRATT, H. K. & WORKMAN, M. Studies on the physiology of tomato fruits. III. The efects of ethylene on respiration and ripening behaviour of fruits stored at 20oC after harvest. Proc. Hort. Sci., n.81, p467478, 1962.
ROMBALDI C.V., BELFAIZA M., RAYNALD J., LATCHE A., PECH J.C. 1-aminocyclopropane-1carboxylate oxidase in ripening fruits: cellular and tissular localization. In: International Symposium on Postharvest Physiology, Pathology and Technologies for Horticultural Commodities, 1994, Agadir, Marrocos, 1994, p35-39.
SCHUCH, W.; KANCZLEER, J.; ROBERTSON, D.; HOBSON, G.; TUCKER G.; GRIERSON, D.; BRIGHT, S.; BIRD, C. Fruit quality characteristics of transgenic tomato fruit with altered polygalacturonase activity. Hort Science, n. 12, p1517-1520, 1991.
SLATER A., MAUNDERS M. J., EDWARDS K., SCHUCH W., GRIERSON D. Isolation and characterization of cDNA clones for tomato polygalactorunase and other ripening related proteins. Plant. Mol. Biol., n. 5, p137-147, 1986.
THEOLOGIS A. What a gas ! Current Biology, n. 3, p369-3371, 1993.
VERVERIDIS P. & JOHN P. Complete recovery in vitro of ethylene-forming enzyme activity. Phytochemistry, n. 30, p725-727, 1991.
WANG H. & WOODSON W.R. A flower senescencerelated mRNA from carnation shares sequence similarity with fruit ripening-related mRNAs involved in ethylene biosynthesis. Plant Physiol., n. 96, p10001001, 1991.
YANG S. F. & HOFFMAN N. E. Ethylene biosynthesis and its regulation in higher plants. Annu. Rev. Plant Physiol., n. 35, p155-189. 1984.
ROMBALDI, Cesar V. 1 ; CHAVES, Ana L. S. 1 ; SILVA, Jorge A. 1; LEMOS, Miriam R. B.; AYUB, Ricardo A.2;
BALAGUÉ, Claudine3; PECH, Jean-Claude3
ctajorge[arroba]brturbo.com.br
1 UFPEL/FAEM - Depto. Ciência e Tecnologia Agroindustrial - Campus Universitário - Caixa Postal, 354 - CEP 96010
900 - Tel. (0532) 75 7258 - Pelotas / RS - Brasil.
2UEPG/FA - Depto. Fitotecnia - Campus Universitário - Ponta Grossa / PR - Brasil.
3INPT / ENSAT - Laboratoire Éthylène et Maturation des Fruits - 147, Avenue de Muret
31300 Toulouse - França.
(Recebido para publicação em 30/11/95)
| Página anterior | Voltar ao início do trabalho | Página seguinte |
|
|
|