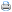Página anterior Página anterior | Voltar ao início do trabalho | Página seguinte  |
O experimento foi realizado no setor de Avicultura do Departamento de Zootecnia da Universidade Federal de Viçosa utilizando-se 400 codornas européias (Coturnix coturnix coturnix), fêmeas, com um dia de idade. As aves foram alojadas e recriadas em piso, onde receberam ração formulada segundo exigências descritas pelo NRC (1994), contendo 24% de PB e 2.900 kcal de EM/kg até o início da produção de ovos.
Aos 40 dias de idade, as aves foram pesadas individualmente. Duzentas e quarenta aves foram selecionadas segundo a conformação e o peso corporal e, posteriormente, foram distribuídas em delineamento inteiramente casualizado, com cinco tratamentos (níveis de energia), seis repetições e oito aves por unidade experimental. As aves foram alojadas em gaiolas de arame galvanizado, de modo que todas as unidades experimentais apresentassem o mesmo peso médio inicial. As gaiolas (1,0 m de comprimento x 0,23 m de largura x 0,20 m de altura) foram dispostas em três andares, montadas em esquema de escada.
Cada gaiola foi subdividida em duas repartições iguais de 0,50 m, com capacidade para oito aves, proporcionando uma área de 115 cm2/ave. Sobre o piso de cimento, logo abaixo das gaiolas, foi disposta uma camada de maravalha para absorção da umidade das excretas. O comedouro foi posicionado na parte frontal e o bebedouro na parte posterior da gaiola, sendo ambos do tipo calha e em chapa galvanizada.
Durante o período pré-experimental (40 a 56 dias de idade), as aves receberam rações contendo 20% de PB, 2.900 kcal de EM/kg e 2,5% de Ca, conforme o NRC (1994). Aos 50 dias de idade, as unidades experimentais foram reavaliadas para que todas apresentassem a mesma produção de ovos. O período experimental foi iniciado aos 56 dias de idade, sendo composto de quatro períodos de 28 dias, totalizando 112 dias.
As exigências nutricionais para a fase de produção (56 a 168 dias) utilizadas nas formulações das dietas foram as preconizadas pelo NRC (1994), exceto as de metionina+ cistina e lisina, que foram baseadas nas recomendações de Pinto et al. (2003a,b).
Para determinação das exigências energéticas, foram formuladas cinco dietas isoprotéicas e isoaminoacídias, à base de milho e farelo de soja, contendo cinco níveis de EM (2.650, 2.750, 2.850, 2.950 e 3.050 kcal de EM/kg), constituindo os tratamentos experimentais (Tabela 1). Essas rações foram formuladas com base nas composições dos ingredientes descritas por Rostagno et al. (2000).
A água foi disponibilizada à vontade e a ração foi fonecida duas vezes ao dia, às 8 e 16h. O programa de iluminação teve início por volta do 40º dia de idade, com fornecimento inicial de 14 horas de luz diária e com aumentos semanais de 30 minutos até atingir 17 horas de luz por dia, programa mantido até o final do período experimental.
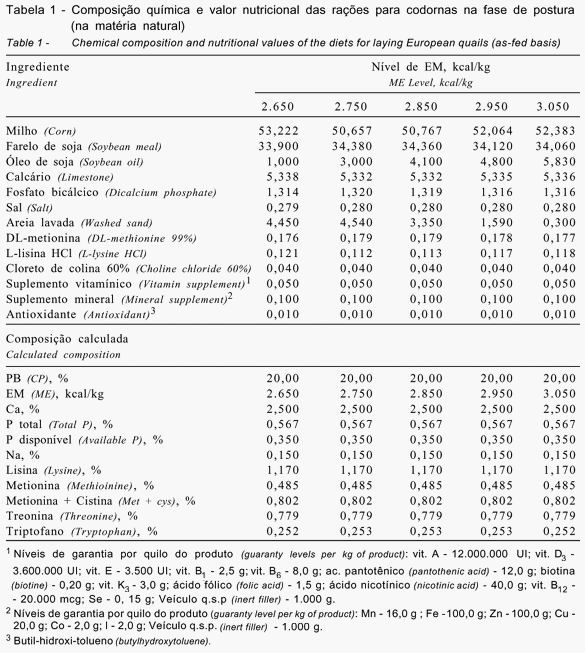
Foram analisados os consumos de ração (g/ave/dia) e de energia (kcal de EM/ave/dia), a produção total de ovos (%/ave/dia) e de ovos comercializáveis (%/ave/dia), o peso (g) e a massa de ovos (g/ave/dia), a conversão alimentar (kg de ração/kg de ovo e kg de ração/dúzia de ovo), o ganho de peso corporal (g), a eficiência energética (kcal de EM/dúzia de ovo e kcal de EM/kg de ovo), os pesos médios de gema, de albúmem e de casca (g) e a concentração de colesterol na gema (mg/g).
A produção de ovos comercializáveis (PROC) refere-se à porcentagem de ovos viáveis à comercialização, excluindo-se os quebrados, trincados ou anormais (DEF), e foi obtida pela fórmula: PROC = 100 - DEF.
Durante os três últimos dias de cada período, foram pesados todos os ovos íntegros produzidos em cada repetição para avaliação do peso médio dos ovos. Para obtenção dos componentes do ovo, foram avaliados os pesos de gema, albúmem e casca utilizando-se quatro ovos de cada repetição, coletados aleatória e diariamente do total de ovos coletados nos três últimos dias de cada período. Os ovos de cada repetição e de cada dia foram pesados individualmente em balança com precisão de 0,01 g e, após as pesagens, foram identificados e quebrados. A gema de cada ovo foi pesada e a respectiva casca foi lavada e seca ao ar para posterior obtenção do peso da casca. O peso do albúmem foi calculado como a diferença entre o peso do ovo e os pesos de gema e casca.
O teor de colesterol na gema foi determinado utilizando-se metodologia proposta por Bragagnolo & Amaya (2003), aplicada por meio do HPLC/RP (Cromatografia Líquida de Alta Performance).
Os parâmetros avaliados foram submetidos à análise de variância a 5% utilizando-se o programa SAEG - Sistema para Análises Estatísticas e Genéticas (UFV, 2004). Posteriormente, os efeitos dos níveis de energia foram estimados por meio de análise de variáveis pelos modelos de regressão linear e quadrática, conforme o melhor ajustamento obtido para cada variável, considerando o comportamento biológico das aves.
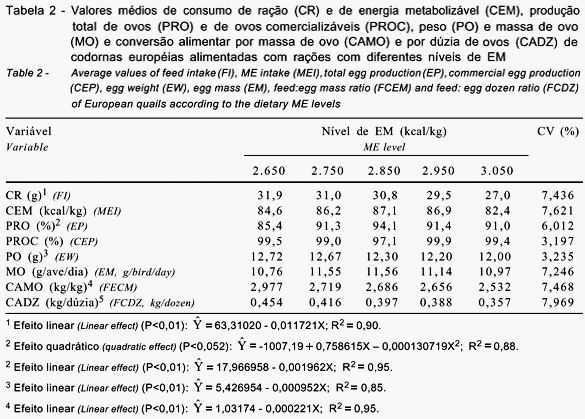
Houve efeito significativo (P<0,01) dos níveis de EM na ração sobre o consumo médio de ração, a produção de ovos, o peso do ovo, a conversão alimentar por dúzia e por massa de ovos (Tabela 2), a eficiência energética por dúzia de ovos e os pesos de gema e de albúmem (Tabela 3). No entanto, os níveis de EM não influenciaram significativamente (P>0,05) o consumo diário de EM, a produção de ovos comercializáveis, a massa de ovo (Tabela 2), a eficiência energética por massa de ovos, o ganho de peso das aves, o peso de casca e a concentração de colesterol na gema do ovo (Tabela 3).
Verificou-se redução linear (P<0,01) diária de 0,01172
g/ave no consumo de ração para cada kcal de aumento no nível
de EM/kg de ração ( ![]() =
63,31020 - 0,011721X; R2 = 0,90). Sabe-se que o nível energético
da dieta constitui fator regulador da ingestão de nutrientes e da EM.
Esta quantidade de EM consumida depende ainda das necessidades da ave, do peso
corporal, da fase de postura, do crescimento, dos níveis de mantença
e do ambiente de criação.
=
63,31020 - 0,011721X; R2 = 0,90). Sabe-se que o nível energético
da dieta constitui fator regulador da ingestão de nutrientes e da EM.
Esta quantidade de EM consumida depende ainda das necessidades da ave, do peso
corporal, da fase de postura, do crescimento, dos níveis de mantença
e do ambiente de criação.
De fato, o consumo diário de energia manteve-se praticamente constante em todos os tratamentos, variando de 82,4 a 87,1 kcal de EM/ave, possivelmente em virtude da redução no consumo com o aumento da energia da ração. Como a variação no consumo de energia foi pequena, este efeito foi não significativo (P>0,05), apesar da grande amplitude entre o menor e o maior nível de EM avaliado (2.650 e 3.050 kcal/kg, respectivamente), indicando que o consumo por codornas, em relação ao nível de energia da ração, é similar ao de galinhas poedeiras (Begin, 1968). Esses resultados corroboram aqueles verificados por Murakami et al. (1993) e Freitas et al. (2005), que também notaram redução linear do consumo de ração com o aumento do nível de energia de 2.500 para 3.100 kcal e de 2.585 a 2.885 kcal/kg de ração, respectivamente. O consumo diário de energia observado neste trabalho está de acordo também com o verificado por Freitas et al. (2005), que constataram não haver efeito significativo no consumo diário de energia de codornas japonesas.
Segundo Yamane et al. (1980) e NRC (1994), o consumo diário de energia por codornas japonesas para máxima produção deve ser de 63 a 75 kcal. O consumo de energia por codornas européias neste experimento variou de 82,4 a 87,1 kcal de EM/ave, dependendo do nível de energia da ração. Oliveira (2003), no entanto, encontrou para codornas européias consumo de 100,5 kcal, em ração contendo 2.800 kcal de EM/kg. Estes dados indicam que as codornas européias, por seu maior peso corporal e metabólico, necessitam de maior aporte energético diário para manutenção de suas atividade vitais.
A produção total de ovos/ave/dia sofreu efeito quadrático
(P<0,052) dos níveis de energia, observando-se maior produção
nas aves alimentadas com ração contendo 2.900 kcal de EM/kg (
![]() =
-1007,19 + 0,758615X - 0,000130719X2; R2 = 0,88). Esses
dados divergem dos obtidos por Yamane et al. (1980), Murakami et al. (1993)
e Cordeiro et al. (2003), que verificaram redução na porcentagem
de postura com o aumento do nível de energia da ração.
Diferem também daqueles registrados por Freitas et al. (2005), Belo et
al. (2000) e Pinto et al. (2002), que não notaram efeito dos níveis
de energia sobre a produção de ovos. Estes autores também
mantiveram o nível fixo de proteína na ração, independentemente
dos níveis de energia avaliados.
=
-1007,19 + 0,758615X - 0,000130719X2; R2 = 0,88). Esses
dados divergem dos obtidos por Yamane et al. (1980), Murakami et al. (1993)
e Cordeiro et al. (2003), que verificaram redução na porcentagem
de postura com o aumento do nível de energia da ração.
Diferem também daqueles registrados por Freitas et al. (2005), Belo et
al. (2000) e Pinto et al. (2002), que não notaram efeito dos níveis
de energia sobre a produção de ovos. Estes autores também
mantiveram o nível fixo de proteína na ração, independentemente
dos níveis de energia avaliados.
De modo geral, a produção de ovos manteve-se bem elevada em todos os tratamentos, variando de 91 a 94%, com exceção do tratamento no qual as aves foram alimentadas com ração contendo 2.650 kcal de EM/kg (84,6%). A produção de ovos obtida com a ração contendo 2.850 kcal de EM/kg foi 9,2; 3,0; 2,9 e 3,3% superior àquela registrada nos tratamentos com 2.650, 2.750, 2.950 e 3.050 kcal de EM/kg, respectivamente. Ressalta-se também que a produção de ovos por esta subespécie não foi inferior à obtida com codornas japonesas na fase inicial de postura (Pinto et al., 2002; Pereira, 2004; Freitas et al., 2005), o que comprova que estas subespécies são igualmente eficientes quanto ao número de ovos produzidos nessa fase. Oliveira (2003) registrou em codornas européias produção média semanal de 6,3 ovos/ave durante as primeiras 41 semanas de produção, o que corresponde a 90% de produção de ovos/ave/dia, indicando também excelente desempenho desta subespécie.
Diferentemente do observado para a produção de ovos/ave/dia, os níveis de energia não afetaram (P>0,05) a produção de ovos comercializáveis, que foi elevada em praticamente todos os tratamentos, não ocorrendo perdas significativas em nenhum dos tratamentos.
Verificou-se redução (P<0,01) de 0,00196 g no peso médio
dos ovos para cada aumento de 1 kcal de EM/kg de ração ( ![]() =
17,966958 - 0,001962X; R2 = 0,95), o que está de acordo com
o observado por Freitas et al. (2005), Belo et al. (2000) e Pinto et al. (2002),
que também notaram redução do peso do ovo com o aumento
do nível energético da ração. A variação
no peso foi de 0,72 g entre os pesos obtidos para o menor e o maior nível
de energia avaliado, provavelmente em virtude do menor consumo de ração
e, conseqüentemente, de todos os nutrientes, principalmente proteína
e metionina, que influenciaram o peso, mas não a produção
de ovos. Sabe-se que, em condições de deficiência de proteína
na ração, o tamanho dos ovos tende a reduzir, enquanto a produção
é mantida com objetivo de perpetuar a espécie. As aves alimentadas
com ração contendo 2.650 kcal de EM/kg de ração
mantiveram consumo diário de 155 mg de metionina, enquanto aquelas que
receberam ração contendo 3.050 kcal de EM/kg consumiram 131 mg
de metionina, sendo este consumo 15,5% inferior.
=
17,966958 - 0,001962X; R2 = 0,95), o que está de acordo com
o observado por Freitas et al. (2005), Belo et al. (2000) e Pinto et al. (2002),
que também notaram redução do peso do ovo com o aumento
do nível energético da ração. A variação
no peso foi de 0,72 g entre os pesos obtidos para o menor e o maior nível
de energia avaliado, provavelmente em virtude do menor consumo de ração
e, conseqüentemente, de todos os nutrientes, principalmente proteína
e metionina, que influenciaram o peso, mas não a produção
de ovos. Sabe-se que, em condições de deficiência de proteína
na ração, o tamanho dos ovos tende a reduzir, enquanto a produção
é mantida com objetivo de perpetuar a espécie. As aves alimentadas
com ração contendo 2.650 kcal de EM/kg de ração
mantiveram consumo diário de 155 mg de metionina, enquanto aquelas que
receberam ração contendo 3.050 kcal de EM/kg consumiram 131 mg
de metionina, sendo este consumo 15,5% inferior.
Com base na composição do ovo e em pesquisas, Begin & Insko Jr. (1972) relataram que codornas japonesas em postura alimentadas com rações à base de milho e farelo de soja requerem 20% de PB quando a ração contém 2.750 kcal de EM/kg e 22% quando o teor de energia é de 2.890 kcal de EM/kg. Os autores recomendaram para obtenção de máxima produção de ovos consumo diário de pelo menos 4,71 g de proteína/ave para manutenção da taxa de postura em torno de 80%. Yamane et al. (1980), Pinto et al. (2002) e Freitas et al. (2005) recomendaram para codornas japonesas consumo diário de 4,9; 6,0 e 4,4 g de proteína/ave, respectivamente, para ótimos índices de produção. Neste estudo, as aves alimentadas com a ração contendo 3.050 kcal de EM/kg apresentaram consumo diário de proteína de 5,4 g, enquanto aquelas alimentadas com ração contendo 2.650 e 2.750 kcal/kg tiveram consumo diário de 6,4 e 6,2 g de proteína/ave, respectivamente. O consumo diário de 5,4 g de PB nas aves alimentadas com ração contendo 3.050 kcal de EM/kg foi 12,9 e 15,6% inferior ao obtido com os níveis de 2.750 de 2.650 kcal/kg, respectivamente, resultando em menor peso do ovo. Pode-se inferir, portanto, que o consumo diário de 6,2 g de proteína/ave parece ser apropriado para a codorna européia. Este valor é próximo ao recomendado por Pinto et al. (2002) para codornas japonesas (6,0 g).
De acordo com Perly (1979), Murakami & Furlan (2002) e Pinto et al. (2002), o peso do ovo é altamente dependente da ingestão diária de proteína em poedeiras, podendo ser utilizado como referência para garantir suas exigências em aminoácidos.
O peso dos ovos destas codornas foi mais alto que os observados por Freitas et al. (2005) na subespécie japonesa, de 10,3 a 10,0 g para o maior e o menor nível de energia, respectivamente, obtidos com os níveis de 2.585 a 2.885 kcal de EM/kg. Pinto et al. (2002) encontraram valores de 10,68 a 10,85 g ao avaliarem níveis de 3.050 a 2.850 kcal de EM/kg de ração. No entanto, trabalhando com quatro grupos genéticos de codornas européias alimentadas com ração contendo 2.900 kcal de EM/kg e 20% de PB, Móri et al. (2005b) encontraram para o peso do ovo variação de 12,81 a 13,45 g. Portanto, desconsiderando a conversão alimentar, o peso do ovo pode ser considerado outra vantagem na criação de codornas européias, principalmente quando se pretende processá-los e vendê-los descascados, por peso, agregando valor ao produto, quando o interesse é a criação e exploração de codornas européias (carne e ovos).
Apesar da redução do peso do ovo e da melhora da produção de ovos, a massa de ovos não foi significativamente (P>0,05) afetada pela elevação dos níveis energéticos da ração. A redução foi de 10,76 e de 10,97 g para o menor e o maior nível de EM, respectivamente, e correspondeu a um pequeno aumento, não-significativo, de 1,95%. Resultados contraditórios, com redução linear de 7,7% na massa diária de ovo, foram observados por Freitas et al. (2005), entre o menor e o maior nível de energia avaliado, de 2.585 e 2.885 kcal de EM/kg de ração, respectivamente.
Em decorrência da redução do consumo de ração,
de 15,4% entre o menor e o maior nível de energia da dieta, e da ausência
de diferenças significativas para a massa dos ovos, houve melhoras significativas
(P<0,01), de 0,00022 pontos na conversão alimentar por dúzia
de ovos ( ![]() =
1,03174 - 0,000221X; R2 = 0,95) e de 0,00095 pontos na conversão
alimentar por massa de ovos (
=
1,03174 - 0,000221X; R2 = 0,95) e de 0,00095 pontos na conversão
alimentar por massa de ovos ( ![]() =
5,426954 - 0,000952X; R2 = 0,85), para cada aumento de 1 kcal de
EM/kg de ração.
=
5,426954 - 0,000952X; R2 = 0,85), para cada aumento de 1 kcal de
EM/kg de ração.
Estes resultados corroboram os encontrados por Pinto et al. (2002), que verificaram que os níveis entre 2.950 e 3.050 kcal de EM/kg permitiram os melhores valores para a conversão alimentar de codornas japonesas. Estão também parcialmente de acordo com os de Murakami et al. (1993), que registraram melhora da conversão alimentar por dúzia de ovos, mas não por massa de ovos, com o aumento de 2.500 para 3.000 kcal de EM/kg nos níveis de energia das rações. Freitas et al. (2005), no entanto, não encontraram diferenças significativas para a conversão alimentar por massa de ovo ao avaliarem rações formuladas com 2.585 a 2.885 kcal de EM/kg.
Ressalta-se que uma das desvantagens na criação da codorna européia, em comparação às codornas japonesas, é seu maior peso (100-120 g a mais) e mais alto consumo de ração (8-12 g a mais), que contribuem para piora na conversão alimentar. Todavia, em estudo com codornas européias, Oliveira (2003) encontrou valores de 0,47 kg/dúzia de ovos e de 3,12 para a conversão alimentar por massa de ovos. Neste trabalho, a conversão alimentar por massa de ovos variou de 2,98 a 2,53 entre os níveis de energia avaliados, enquanto, nos trabalhos de Freitas et al. (2005), com níveis de energia próximos ao deste trabalho, a conversão foi de 3,19 a 3,27.
Verificou-se que apenas a eficiência energética para produção
de uma dúzia de ovos, em comparação à eficiência
para produção de 1 kg de ovos, foi significativamente influenciada
(P<0,01) pelos níveis de EM. Houve redução de 0,2235
kcal na eficiência energética das aves para produção
de uma dúzia de ovos a cada aumento de 1 kcal de EM/kg de ração
( ![]() =
1781,049 - 0,223504X; R2 = 0,77). Esses dados revelam que foi mais
eficiente, energeticamente, produzir uma dúzia de ovos com o fornecimento
de 3.050 kcal de EM/kg de ração. Entretanto, Belo (1997) não
encontraram efeito dos níveis de energia da ração sobre
a eficiência energética para produção de uma dúzia
ou de 1 kg de ovos.
=
1781,049 - 0,223504X; R2 = 0,77). Esses dados revelam que foi mais
eficiente, energeticamente, produzir uma dúzia de ovos com o fornecimento
de 3.050 kcal de EM/kg de ração. Entretanto, Belo (1997) não
encontraram efeito dos níveis de energia da ração sobre
a eficiência energética para produção de uma dúzia
ou de 1 kg de ovos.
Os grupos de aves alimentadas com dietas formuladas com 2.650 a 3.050 kcal de EM/kg não diferiram significativamente (P>0,05) quanto ao ganho e à perda de peso. Esse resultado corrobora os obtidos por Pinto et al. (2002), que também não notaram efeito do nível de energia da ração sobre o ganho de peso das aves no período experimental. Com base nos resultados, pode-se inferir que todas as rações foram eficientes para manutenção do peso, o que poderia afetar a produtividade. As dietas contendo o menor e o maior nível de energia proporcionaram os menores ganhos de peso em valor absoluto. No entanto, o baixo ganho de peso das aves alimentadas com a ração contendo o maior nível de energia (3.050 kcal de EM/kg) foi suficiente para manter elevada produção, quando comparado ao mesmo ganho obtido com a dieta contendo o menor nível de energia (2.650 kcal de EM/kg).
Observou-se redução significativa (P<0,01) de 0,00043 g no
peso de gema ( ![]() =
5,209375 - 0,000429X; R2 = 0,72) e de 0,00130 g no peso de albúmem
(
=
5,209375 - 0,000429X; R2 = 0,72) e de 0,00130 g no peso de albúmem
( ![]() =
11,182973 - 0,001302X; R2 = 0,91) com o aumento do nível de
energia da ração. Entretanto, o mesmo não foi verificado
para o peso da casca do ovo. A redução no peso da gema e no peso
de albúmem foi acompanhada pela redução no peso do ovo
com o aumento do nível de EM da ração, provavelmente em
virtude da menor quantidade de nutrientes necessários à formação
do ovo. Fonseca (1992), em experimento com galinhas poedeiras, forneceu ração
contendo níveis mais baixos de energia e obteve ovos com maior espessura
de casca.
=
11,182973 - 0,001302X; R2 = 0,91) com o aumento do nível de
energia da ração. Entretanto, o mesmo não foi verificado
para o peso da casca do ovo. A redução no peso da gema e no peso
de albúmem foi acompanhada pela redução no peso do ovo
com o aumento do nível de EM da ração, provavelmente em
virtude da menor quantidade de nutrientes necessários à formação
do ovo. Fonseca (1992), em experimento com galinhas poedeiras, forneceu ração
contendo níveis mais baixos de energia e obteve ovos com maior espessura
de casca.
O teor de colesterol na gema do ovo não foi influenciado (P>0,05) pelo nível de energia da ração, o que confirma os achados de Quirino et al. (2004), que também não notaram aumento no teor de colesterol na gema dos ovos de codornas japonesas alimentadas com ração contendo os mesmos níveis de energia avaliados neste trabalho.
Rações para codornas européias em fase inicial de postura (56 aos 168 dias) devem conter 3.050 kcal de EM/kg (correspondente a um consumo de 82,4 kcal de EM/ave/dia ou a 7,5 kcal de EM/g de ovo) para obtenção de melhor conversão alimentar por massa e por dúzia de ovos ou 2.900 kcal de EM/kg (correspondente a um consumo de 87 kcal de EM/ave/dia ou a 7,5 kcal de EM/g de ovo) para melhor produção de ovos.
ALMEIDA, M.I.M. Efeito de linhagem e de nível protéico sobre o desempenho e características de carcaça de codornas (Coturnix sp) criadas para corte. Botucatu: Universidade Estadual Paulista, 2001. 96p. Tese (Doutorado em Melhoramento Genético) - Universidade Estadual Paulista, 2001.
ALBINO, L.F.T.; BARRETO, S.L.T. Criação de codornas para produção de ovos e carnes. Viçosa, MG: Aprenda Fácil, 2003. 289p.
BELO, M.T.S. Níveis de energia metabolizável e de metionina em rações de codornas japonesas (Coturnix coturnix japonica) na fase inicial de postura. Lavras: Universidade Federal de Lavras, 1997. 60p. Dissertação (Mestrado em Zootecnia) - Universidade Federal de Lavras, 1997.
BELO, M.T.S.; COTTA, J.T.B.; OLIVEIRA, A.I.G. et al. Níveis de energia metabolizável em rações de codornas japonesas (Coturnix coturnix japonica) na fase inicial de postura. Ciências Agrotécnicas, v.24, n.3, p.782-794, 2000.
BEGIN, J.J. A comparison of the ability of the Japanese quail and light breed chicken to metabolizable and utilize energy. Poultry Science, v.47, n.4, p.1278-1281, 1968.
BEGIN, J.J.; INSKO JR., W.M. The Effects of dietary protein level on the reproductive performance of coturnix breeder hens. Poultry Science, v.51, n.5, p.1662-1668, 1972.
BRAGAGNOLO, N.R.; AMAYA, D.B. Comparison of the cholesterol content of Brazilian chicken and quail eggs. Journal of Food Composition and Analysis, v.16, p.147-153, 2003.
CORDEIRO, M.D.; SOARES, R.T.R.N.; AVILA, R.P. et al. Níveis de energia metabolizável para codornas japonesas (Coturnix coturnix japonica) na fase inicial de postura. In: REUNIÃO ANUAL DA SOCIEDADE BRASILEIRA DE ZOOTECNIA, 40., 2003, Santa Maria. Anais... Santa Maria: Sociedade Brasileira de Zootecnia, 2003. (CD-ROM).
FREITAS, A.C.; FUENTES, M.F.F.; FREITAS, E.R. et al. Efeito de níveis de proteína bruta e de energia metabolizável na dieta sobre o desempenho de codornas de postura. Revista Brasileira de Zootecnia, v.34, n.3, p.838-846, 2005.
FONSECA, R.A. Níveis de energia em duas linhagens de poedeiras leves, na fase inicial de postura. Lavras: Universidade Federal de Lavras, 1992. 123p. Dissertação (Mestrado em Zootecnia) - Universidade Federal de Lavras, 1992.
MÓRI, C.; GARCIA, E.A.; PAVAN, A.C. et al. Desempenho e rendimento de carcaça de quatro grupos genéticos de codornas para produção de carne. Revista Brasileira de Zootecnia, v.34, n.3, p.870-876, 2005a.
MÓRI, C.; GARCIA, E.A.; PAVAN, A.C. et al. Desempenho e qualidade dos ovos de codornas de quatro grupos genéticos. Revista Brasileira de Zootecnia, v.34, n.3, p.864-869, 2005b.
MURAKAMI, A.E.; FURLAN, A.C. Pesquisa na nutrição e alimentação de codornas em postura no Brasil. In: SIMPÓSIO INTERNACIONAL DE COTURNICULTURA, 1., 2002, Lavras. Anais... Lavras: Universidade Federal de Lavras, 2002. p.113-120.
MURAKAMI, A.E.; MORAES, V.M.B.; ARIKI, J. Níveis de proteína e energia em dietas de codornas japonesas (Coturnix coturnix japonica) em postura. Revista Brasileira de Zootecnia, v.22, n.4, p.541-551, 1993.
NATIONAL RESEARCH COUNCIL - NRC. Nutrient requirements of poultry. Washington, D.C.: 1994. p.44-45.
NETO, G.J. Aspectos nutricionais que afetam as características especificas do ovo de incubação. In: CONFERÊNCIA APINCO DE CIÊNCIA E TECNOLOGIA AVÍCOLAS, 1999, Campinas. Anais... Campinas: Fundação Apinco de Ciência e Tecnologia Avícolas, 1999. p.145-164.
OLIVEIRA, L.Q.M. Parâmetros produtivos e níveis nutricionais de cálcio para codorna européia na fase de postura. Brasília: Universidade de Brasília, 2003. 55p. Dissertação (Mestrado em Ciências Agrárias) - Universidade de Brasília, 2003.
PEREIRA, C.A. Exigência nutricional de cálcio para codornas japonesas durante o pico de postura. Viçosa, MG: Universidade Federal de Viçosa, 2004. 60p. Dissertação (Mestrado em Zootecnia) - Universidade Federal de Viçosa, 2004.
PERLY, L. Correlação entre índice morfológico, peso do ovo e peso vivo ao final da fase de crescimento em codornas doméstica (Coturnix coturnix japonica). Revista do Setor de Ciências Agrárias, v.1, n.1, p.41-53, 1979.
PINTO, R.; FERREIRA, A.S.; ALBINO, L.F.T. et al. Níveis de proteína e energia para codornas japonesas em postura. Revista Brasileira de Zootecnia, v.31, n.4, p.1761-1770, 2002.
PINTO, R.; DONZELE, J.L.; FERREIRA, A.S. et al. Exigência de metionina mais cistina para codornas japonesas em postura. Revista Brasileira de Zootecnia, v.32, n.5, p.1166-1173, 2003a.
PINTO, R.; FERREIRA, A.S.; DONZELE, J.L. et al. Exigência de lisina para codornas japonesas em postura. Revista Brasileira de Zootecnia, v.32, n.5, p.1182-1189, 2003b.
QUIRINO, B.J.S.; BARRETO, S.L.T.; UMIGI, R.T. et al. Efeitos de níveis de energia para codornas japonesas durante o pico de postura. In: CONGRESSO BRASILEIRO DE COTURNICULTURA, 1., 2004, Lavras. Anais... Lavras: Universidade Federal de Lavras, 2004. p.207.
ROSTAGNO, H.S.; ALBINO, L.F.T.; DONZELE, J.L. et al. Composição de alimentos e exigências nutricionais de aves e suínos (Tabelas brasileiras). Viçosa, MG. Universidade Federal de Viçosa, 2000. 141p.
UNIVERSIDADE FEDERAL DE VIÇOSA - UFV. SAEG - Sistema de análises estatísticas e genéticas. Versão 9,0. Viçosa, MG: Fundação Arthur Bernardes, 2004. (CD-ROM).
YAMANE, T.; ONO, K.; TANAKA, T. Energy requirement of laying Japanese quail. British Poultry Science, v.21, n.6, p.451-455, 1980.
Sergio Luiz de Toledo BarretoI;
Bárbara Josefina de Sousa QuirinoII;
Claudson Oliveira BritoII;
Regina Tie UmigiII;
Marcelle Santana de AraujoII;
Tatiana Cristina da RochaII;
Claiton Gonçalves PereiraIII
IDepartamento de Zootecnia/UFV
IIPós-graduação em Zootecnia DZO/UFV
IIIGraduação em Medicina Veterinária
Revista Brasileira de Zootecnia v.36 n.1 Viçosa jan./fev. 2007
| Página anterior | Voltar ao início do trabalho | Página seguinte |
|
|
|