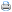Página anterior Página anterior | Voltar ao início do trabalho | Página seguinte  |
O experimento foi desenvolvido no Setor de Ovinocultura, Departamento de Zootecnia da UFLA. Utilizaram-se 36 cordeiros machos inteiros com peso inicial de 15 kg , sendo 12 da raça Bergamácia e 24 da raça Santa Inês, mantidos sob regime de confinamento em gaiolas individuais.
A dieta total utilizada foi constituída de feno triturado de coast cross (Cynodon dactylon (L).Pers.), farelo de soja (Glicine max (L.) Merrill), grão de milho (Zea mays L.), calcário calcítico, suplemento mineral e vitamínico, calculada com base nas exigências nutricionais estabelecidas pelo ARC (1980), para um ganho de peso diário esperado de 300 g. A composição química dos ingredientes utilizados para formulação da dieta e a composição química da mesma são respectivamente apresentadas nas Tabelas 1 e 2.
3 Energia metabolizável determinada para ração total.
Os cordeiros foram sorteados para serem sacrificados a diferentes pesos: 15 kg, 25 kg, 35 kg e 45 kg, resultando para cada peso 03 da raça Bergamácia e 06 da raça Santa Inês. O abate foi feito por meio de um corte na artéria carótida e nas veias jugulares, com coleta e pesagem do sangue.
O período experimental não teve duração préfixada, pois correspondeu ao período necessário para que o último cordeiro atingisse o peso vivo de 45 kg.
Após sangria e retirada da pele, foi feita um abertura ao longo de toda a extensão da linha mediana ventral do abdome. Posteriormente, pesouse o trato gastrointestinal, bexiga e vesícula biliar, cheios e vazios, para determinação do peso corporal vazio (PCVZ). Após a evisceração, retirada da cabeça, pés, cauda e testículos, obteve-se a carcaça do animal.
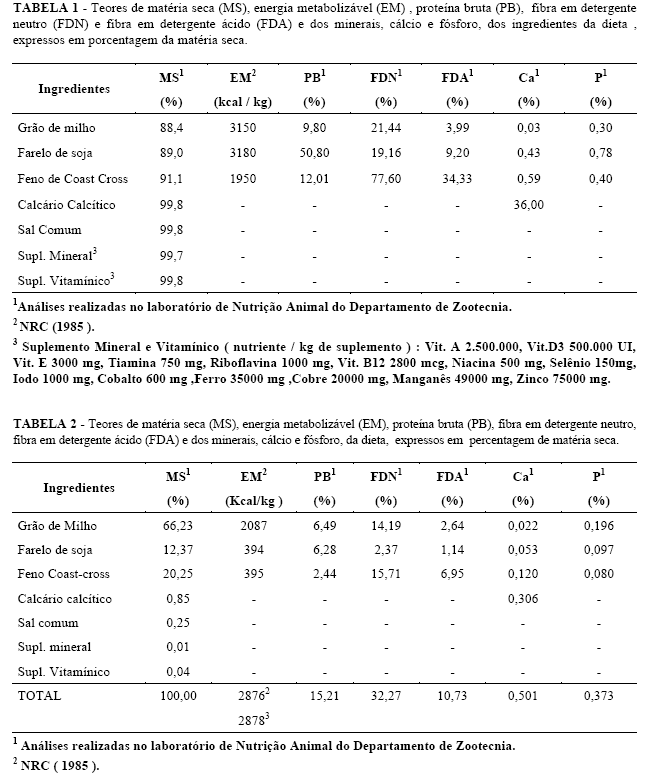
A carcaça quente foi levada à câmara fria a uma temperatura de 2º C, por um período de 24 horas. As carcaças foram mantidas penduradas pela articulação tarso metatarsiana, em ganchos próprios, com distanciamento de 17 cm. Após esse período, procedeuse à retirada do pescoço e, em seguida, mediante corte longitudinal na carcaça, obtiveram-se metades aproximadamente simétricas, pesando-se a ½ carcaça esquerda, que foi dividida em cinco cortes comerciais, perna, lombo, costeleta, costela/fralda e paleta (Figura 1), que tiveram a seguinte base anatônica:
Perna: compreendeu a região sacral e os seguimentos anatômicos do membro pélvico: cíngulo pélvico e perna. Seccionou-se ao nível da articulação da última vértebra lombar e primeira sacral e ao nível da posição média dos ossos do tarso.
Obteve-se por um corte transversal que passou entre a articulação da última vértebra lombar e a primeira sacral, seccionando o ligamento superespinhoso lombo sacro, ligamento interespinhoso e ligamento longitudinal ventral e dorsal, terminando aproximadamente na aponeurose que une o músculo recto do abdome (M. rectus abdominis) com a porção carnosa do oblíquo interno do abdome (M. obliquus internus) .
Lombo: A base óssea desse corte compreendeu da primeira à última (pode ter seis ou sete vértebras) vértebra lombar. Procedeu-se um corte entre a última vértebra torácica e a primeira lombar e outro entre a última lombar e a primeira sacral.
Costeleta: Compreende a região localizada entre a 1ª a 13º vértebra torácica, junto com a metade dorsal do corpo das costelas correspondentes.
Costela / Fralda : esse corte compreendeu a região anatômica da parede abdominal e metade ventral da torácica. Sua base óssea foi metade correspondente do esterno cortado sagitalmente, a metade ventral das oito primeiras costelas e terço ventral das cinco restantes. Obteve-se fazendo-se um corte aproximadamente paralelo à coluna vertebral, partindo desde a prega inguinal e acabando no cordão testicular .Termina no vértice da cartilagem do manubrio esternal ou na articulação da primeira costela com a primeira esternebra.

Paleta: As regiões anatômicas que compreenderam esse corte foram o cíngulo escapular e braço antebraço. A base óssea foi formada pela escápula, úmero, rádio e ossos do carpo. Obteve-se mediante secção da região axilar, dos músculos que unem a escápula e o úmero na parte ventral do tórax. Depois contornou-se a escápula, seccionando os músculos braquiocefálico, omo transversal, trapézio cervical e serrato cervical pela parte superior, e trapézio torácico e rombóides, pela parte posterior do tronco.
As análises para obtenção dos coeficientes alométricos foram realizadas por meio do Statistical Analysis System (SAS, 1996). Para verificação da hipótese b=1 foi realizado o teste "t" (Student) (a = desenvolvimento de "X" e "Y" foram semelhantes no 0,05 e a = 0,01). Se b = 1, o crescimento foi intervalo de denominado isogônico, indicando que as taxas de desenvolvimento de "X" e "Y" foram semelhantes no intervalo de crescimento considerado. Quando b ¹ 1, o crescimento foi chamado heterogônico, sendo positivo (b > 1), órgão de desenvolvimento tardio; ou negativo (b < 1), órgão de desenvolvimento precoce.
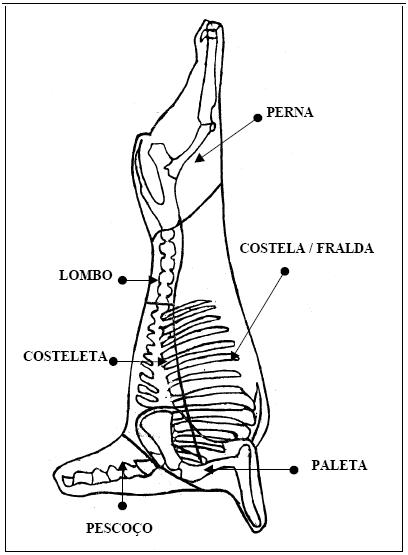
FIGURA 1 - Cortes efetuados na ½ carcaça esquerda de cordeiros Santa Inês e Bergamácia
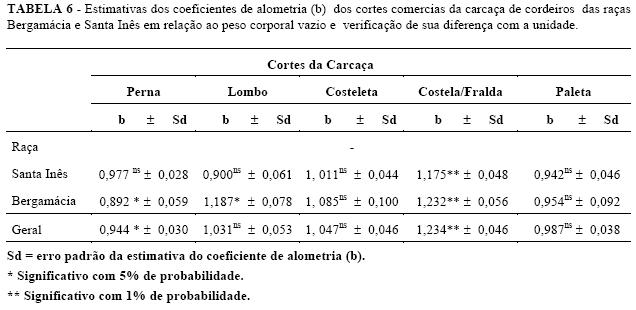
A partir do peso corporal vazio foram determinadas equações de regressão, para estimar o desenvolvimento dos cortes da carcaça: perna, lombo, costeleta, costela/fralda e paleta em relação ao peso corporal vazio de cordeiros Santa Inês (Tabela 3) e Bergamácia (Tabela 4) . Observando a significância dos coeficientes de alometria encontrados para os cortes comerciais, notou-se que o desenvolvimento cordeiros Santa Inês não foram diferentes de um da costeleta, costela/ fralda e paleta foi semelhante (b=1) e para a costela / fralda, segundo o teste de para as duas raças. Os coeficientes alométricos "t", o coeficiente de alometria diferiu de um (b ≠ 1) para perna, lombo, costeleta e paleta dos (Tabela 3).

Na Figura 2 são apresentadas as equações alométricas de cada corte e o comportamento das respectivas linhas de regressão, podendo-se observar que a perna, lombo, costeleta e paleta, crescem sempre acompanhando o desenvolvimento do corpo; no entanto a costela/fralda teve um ritmo de crescimento mais rápido, a partir dos 35 kg de peso corporal vazio dos cordeiros Santa Inês.
Para os cordeiros Bergamácia, os coeficientes alométricos da perna e lombo foram diferentes de um (b 1), o que revelou um desenvolvimento precoce para a perna e tardio (b > 1) para o lombo, em relação ao peso corporal vazio (Tabela 4).
A perna, lombo, costeleta e paleta dos cordeiros Santa Inês, acompanharam o desenvolvimento do corpo, e o mesmo ocorreu com os Bergamácia, quanto ao crescimento da costeleta e da paleta.
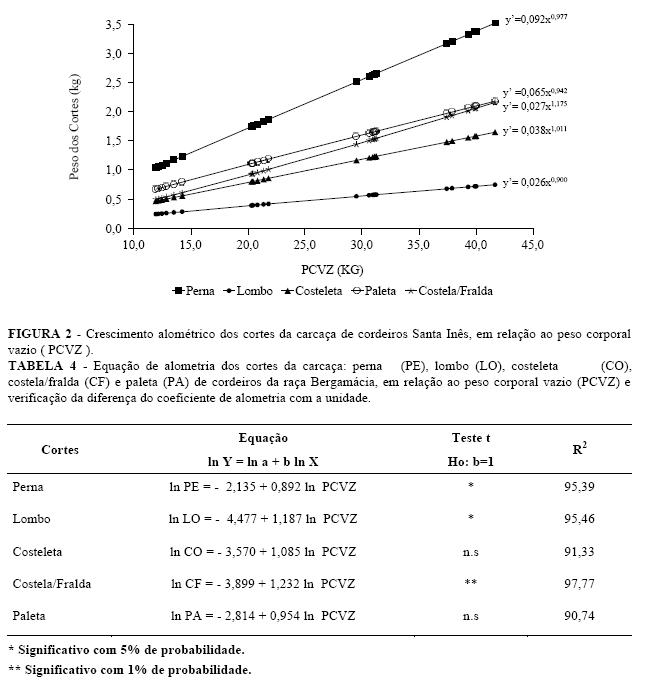
Na Figura 3 são apresentadas as equações alométricas de cada corte e o comportamento das respectivas linhas de regressão, podendo-se observar que o lombo e costela/fralda, apresentaram desenvolvimento relativo tardio, enquanto a perna cresceu de forma precoce e a costeleta acompanhou o desenvolvimento do corpo vazio dos cordeiros Bergamácia.
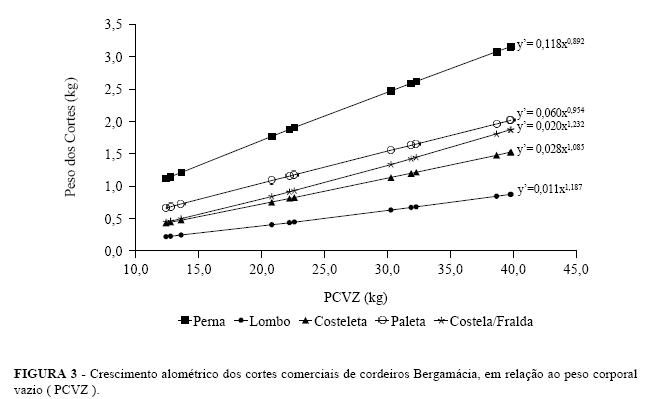
Avaliando o desenvolvimento dos cortes de todos linhas de regressão. De forma geral observou-se que o os cordeiros, observou-se que o lombo, a costeleta e a lombo, a costeleta e a paleta tiveram um crescimento paleta tiveram um desenvolvimento relativo semelhante isogônico em relação ao peso corporal vazio, ou seja, o ao desenvolvimento do corpo vazio, enquanto a perna crescimento desses cortes comerciais foi semelhante ao se desenvolveu de forma precoce e a costela/ fralda, de desenvolvimento relativo do corpo dos cordeiros. forma tardia (Tabela 5). Na Tabela 6 são apresentados os coeficientes
A Figura 4 são apresentadas as equações de alometria dos cortes da carcaça de cordeiros alométricas de cada corte, referente a todos os animais Santa Inês e Bergamácia com seus respectivos erros experimentais e o comportamento das respectivas padrão.

* Significativo com 5% de probabilidade. ** Significativo com 1% de probabilidade.
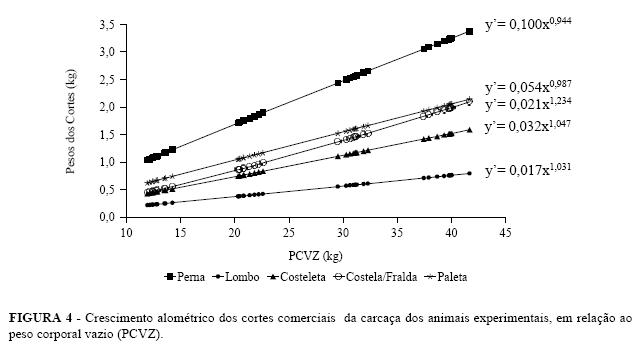
Os coeficientes alométricos para a costeleta (b = 1,011 ± 0,044 e b = 1,085 ± 0,100) e paleta (b = 0,942 ± 0,046 e b = 0,954 ± 0,092) da carcaça dos cordeiros Santa Inês e Bergamácia, respectivamente, demonstram que houve um crescimento isogônico (b =1). Isso indica que a participação relativa do crescimento dessas partes foi semelhante no desenvolvimento do corpo vazio. Todavia, Lohose, Moss e Butterfield (1971) comprovaram um desenvolvimento precoce da paleta e da perna, trabalhando com a raça Ideal. Por outro lado, Roque et al. (1998) trabalhando com as raças Corriedale e Rommey Marsh, verificaram um crescimento isogônico para a região da paleta e da perna, em relação ao peso da carcaça fria. Cabrero (1984), citado por Osório et al. (1995), num experimento com ovinos da raça Merina, verificou que a paleta foi uma das primeiras porções a se desenvolver. Silva (1999) , verificou que em relação a peso do corpo vazio, apenas para a paleta o coeficiente de alometria é menor do que um. Isto indicaria que a paleta cresceria proporcionalmente menos que os demais componentes do corpo vazio, evidenciando um desenvolvimento mais precoce, o que não foi observado neste experimento.
A costela/fralda das duas raças, Santa Inês e Bergamácia , apresentaram, respectivamente, os seguintes coeficientes de alometria: b = 1,175 ± 0,048 e b = 1,232 ± 0,056 , o que implica dizer que o crescimento desse corte foi heterogônico positivo (b > 1) ou seja, a taxa de desenvolvimento foi tardia em relação ao aumento do peso corporal dos cordeiros. O desenvolvimento pode ser mais ou menos tardio ou, em alguns casos, até mesmo isogônico (b = 1), de acordo com o grupo genético, o que pode ser comprovado com os dados obtidos por Roque et al. (1998) que, com relação ao costilhar, apresentou-se tardio na raça Romney Marsh, mas na Corriedale foi isogônico. Osório et al. (1995), trabalhando com cordeiros machos castrados e fêmeas, da raça Ideal e cruzas de ovelhas Ideal x Texel, observaram em relação ao peso da carcaça fria, ter o costilhar apresentado desenvolvimento rápido e heterogônicos para fêmeas e machos, b = 1,179 ± 0,049 e b= 1,416 ± 0,115, respectivamente. Lopez (1987) citado por Roque et al. (1998), verificou que a região do costilhar apresentou desenvolvimento mais tardio em fêmeas que nos machos procedentes de cruzamento. O desenvolvimento tardio da costela/fralda está relacionado ao fato de esse paleta ter sido semelhante ao peso corporal vazio, corte possuir em sua composição uma quantidade considerando que esses cortes possuem mais tecido elevada de gordura. Como a porção muscular constitui muscular que adiposo, o que os difere da costela/ fralda. uma proporção muito constante do peso corporal de O autor ainda ratifica que há uma variação considerável animais com o trato gastrointestinal vazio (Prescott, no conteúdo de gordura à medida que o peso corporal 1982), provavelmente isso explique o fato de o vazio se eleva. desenvolvimento relativo da perna, lombo, costela e paleta ter sido semelhante ao peso corporal vazio, considerando que esses cortes possuem mais tecido muscular que adiposo, o que os difere da costela/ fralda. O autor ainda ratifica que há uma variação considerável no conteúdo de gordura à medida que o peso corporal vazio se eleva.
Considerando as condições em que o experimento foi desenvolvido, os resultados obtidos permitiram estabelecer as seguintes conclusões :
Nenhum dos cortes dos cordeiros Santa Inês apresentou crescimento heterogônico negativo, o crescimento da perna, lombo, costeleta e paleta foi isogônico e da costela/fralda foi heterogônico positivo.
Os cordeiros Santa Inês e Bergamácia se igualam quanto ao desenvolvimento da costeleta e da paleta, o qual relativamente acompanhou o desenvolvimento do corpo vazio, no entanto em relação ao desenvolvimento da perna e do lombo, as raças foram diferentes.
AGRICULTURAL RESEARCH COUNCIL. The nutrient requeriments of farm livestock. London:ARC, 1980. 351p.
ÁVILA, V.; OSÓRIO, J.C.S. Efeito do sistema de criação, época de nascimento e ano na velocidade de crescimento de cordeiros. Revista da Sociedade Brasileira de Zootecnia, Viçosa, v.25, n.5,p.10071016, set./out. 1996.
BERG, R.J.; BUTTERFIELD, R.M. A classification of bovine muscles based on their relative growth patterns. Review Veterinary Science, v.7, p.326332, Sept. 1966.
BERG, R.J.; BUTTERFIELD, R.M. New concepts of cattle growth. Sidney: Sidney University Press, 1976. 240p.
CALLOW, E.A. Comparative studies of meat. II. Changes in carcass during growth and fatting and their relation to chemical composition of fatty muscular tissues. Journal of Agricultural Science, Cambridge, v.38, p.174-199, Dec.1948.
FOWLER, V.R. Body development and some problems of its evaluation. In: LOGDE, G.A .; LANMING, G.E. (eds). Growth and development of mammals. London: Butterworth, 1967. p.105-131.
GOUVEA, R.C.D. Aprenda a criar ovelhas. São Paulo: Três, 1987. 95p.
HUXLEY, J.S. Problems of relative growth. London:Methuen, 1932. 577p.
LOHOSE, C.L.; MOSS, F.P.; BUTTERFIELD, R.M. Growth patterns of muscle of Merino sheep from birth to 517 days. Animal Production, Scotland, v.13, p.117-126., Aug. 1971.
NUNES, J.F.; CIRÍACO, A L.T.; SUASSUNA, U. Produção e reprodução de caprinos e ovinos. 2.ed. Fortaleza: LCR, 1997. 160p.
OSÓRIO, J.C. da S.; SIEWERDT, F.; OSÓRIO, M.T.M. et al. Desenvolvimento alométrico das regiões corporais em ovinos. Revista da Sociedade Brasileira de Zootecnia, Viçosa, v.24, n.2, p.326333, mar./abr.1995.
PRESCOTT, J. H. D. Crecimiento y desarrollo de los corderos. In: HAPEZ, E.S.E. Dessarrolo y nutricion animal. Zaragoza:Acribia, 1982. p.351-369.
ROQUE, A.P.; OSÓRIO, J.C.S.; JARDIM, P.O et al. Desenvolvimento relativo da composição regional e tecidual em cordeiros de cinco raças. In: REUNIÃO ANUAL DA SOCIEDADE BRASILEIRA DE ZOOTECNIA, 1998, Botucatu. Anais... Botucatu: SBZ, 1998. v.3, p.627-629.
SAS INSTITUTE. Users Guide: Statistical Analysis System Institute. 5.ed. North Carolina: Cray, 1996. 956p.
SILVA, L.F. da. Crescimento, composição corporal e exigências nutricionais de cordeiros abatidos com diferentes pesos. Santa Maria:UFSM, 1999. 65p. (Dissertação - Mestrado em Produção Animal).
Cristiane Leal Dos Santos 1 Juan Ramón Olalquiaga Pérez 2 Luciana Castro Geraseev 3 Osni Vieira Prado 3 Joel Augusto Muniz 4
lgeraseev[arroba]nca.ufmg.br
| Página anterior | Voltar ao início do trabalho | Página seguinte |
|
|
|